LaeA, a regulator of secondary metabolism in Aspergillus spp
3
2004
... id="C6">全局性调控是指由全局性调控因子参与并对多个次级代谢物生物合成以及形态分化进行的调控.在诱导丝状真菌产生次级代谢物时,外界环境刺激是一个很重要的因素,而全局性调控因子在其中起着关键作用.LaeA是近年来发现的一种丝状真菌全局性调控因子.2004年,美国威斯康星大学Keller研究组首次在构巢曲霉(Aspergillus nidulans)中分离到laeA基因,生物信息分析表明LaeA含有一个保守的参与组蛋白甲基化的S-腺苷甲硫氨酸(SAM)结合结构域[1],同时发现laeA的启动子区含有途径特异性调控蛋白AflR的结合位点.随后laeA在烟曲霉(A. funigatus)、黄曲霉(A. flavus)、米曲霉(A. oryzae)、黑曲霉(A. niger)、产黄青霉、草酸青霉(P. oxalicum)、橘青霉(P. citrinum)、里氏木霉(Trichodermareesei)、深绿木霉(T. atroviride)、丛毛红曲霉(Monascuspilosus)、红色红曲霉(M. ruber)、灰葡萄孢菌(Botrytis cinerea)、藤仓镰孢菌(Gibberella fujikuroi)、轮枝镰刀菌(Fusarium verticillioide)、禾旋孢腔菌(Cochliobolus sativus)和玉米小斑病菌(Helminthosporium maydis)中被相继报道[2].研究发现laeA能够正调控包括抗生素和真菌毒素在内的大量次级代谢物的生物合成,同时影响真菌的形态分化和发育过程[3].LaeA还能够通过刺激沉默基因的表达激活隐性代谢物的产生[4]. ...
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
... [1]. ...
丝状真菌全局性调控因子LaeA的研究进展
1
2017
... id="C6">全局性调控是指由全局性调控因子参与并对多个次级代谢物生物合成以及形态分化进行的调控.在诱导丝状真菌产生次级代谢物时,外界环境刺激是一个很重要的因素,而全局性调控因子在其中起着关键作用.LaeA是近年来发现的一种丝状真菌全局性调控因子.2004年,美国威斯康星大学Keller研究组首次在构巢曲霉(Aspergillus nidulans)中分离到laeA基因,生物信息分析表明LaeA含有一个保守的参与组蛋白甲基化的S-腺苷甲硫氨酸(SAM)结合结构域[1],同时发现laeA的启动子区含有途径特异性调控蛋白AflR的结合位点.随后laeA在烟曲霉(A. funigatus)、黄曲霉(A. flavus)、米曲霉(A. oryzae)、黑曲霉(A. niger)、产黄青霉、草酸青霉(P. oxalicum)、橘青霉(P. citrinum)、里氏木霉(Trichodermareesei)、深绿木霉(T. atroviride)、丛毛红曲霉(Monascuspilosus)、红色红曲霉(M. ruber)、灰葡萄孢菌(Botrytis cinerea)、藤仓镰孢菌(Gibberella fujikuroi)、轮枝镰刀菌(Fusarium verticillioide)、禾旋孢腔菌(Cochliobolus sativus)和玉米小斑病菌(Helminthosporium maydis)中被相继报道[2].研究发现laeA能够正调控包括抗生素和真菌毒素在内的大量次级代谢物的生物合成,同时影响真菌的形态分化和发育过程[3].LaeA还能够通过刺激沉默基因的表达激活隐性代谢物的产生[4]. ...
丝状真菌全局性调控因子LaeA的研究进展
1
2017
... id="C6">全局性调控是指由全局性调控因子参与并对多个次级代谢物生物合成以及形态分化进行的调控.在诱导丝状真菌产生次级代谢物时,外界环境刺激是一个很重要的因素,而全局性调控因子在其中起着关键作用.LaeA是近年来发现的一种丝状真菌全局性调控因子.2004年,美国威斯康星大学Keller研究组首次在构巢曲霉(Aspergillus nidulans)中分离到laeA基因,生物信息分析表明LaeA含有一个保守的参与组蛋白甲基化的S-腺苷甲硫氨酸(SAM)结合结构域[1],同时发现laeA的启动子区含有途径特异性调控蛋白AflR的结合位点.随后laeA在烟曲霉(A. funigatus)、黄曲霉(A. flavus)、米曲霉(A. oryzae)、黑曲霉(A. niger)、产黄青霉、草酸青霉(P. oxalicum)、橘青霉(P. citrinum)、里氏木霉(Trichodermareesei)、深绿木霉(T. atroviride)、丛毛红曲霉(Monascuspilosus)、红色红曲霉(M. ruber)、灰葡萄孢菌(Botrytis cinerea)、藤仓镰孢菌(Gibberella fujikuroi)、轮枝镰刀菌(Fusarium verticillioide)、禾旋孢腔菌(Cochliobolus sativus)和玉米小斑病菌(Helminthosporium maydis)中被相继报道[2].研究发现laeA能够正调控包括抗生素和真菌毒素在内的大量次级代谢物的生物合成,同时影响真菌的形态分化和发育过程[3].LaeA还能够通过刺激沉默基因的表达激活隐性代谢物的产生[4]. ...
LaeA, a regulator of morphogenetic fungal virulence factors
1
2005
... id="C6">全局性调控是指由全局性调控因子参与并对多个次级代谢物生物合成以及形态分化进行的调控.在诱导丝状真菌产生次级代谢物时,外界环境刺激是一个很重要的因素,而全局性调控因子在其中起着关键作用.LaeA是近年来发现的一种丝状真菌全局性调控因子.2004年,美国威斯康星大学Keller研究组首次在构巢曲霉(Aspergillus nidulans)中分离到laeA基因,生物信息分析表明LaeA含有一个保守的参与组蛋白甲基化的S-腺苷甲硫氨酸(SAM)结合结构域[1],同时发现laeA的启动子区含有途径特异性调控蛋白AflR的结合位点.随后laeA在烟曲霉(A. funigatus)、黄曲霉(A. flavus)、米曲霉(A. oryzae)、黑曲霉(A. niger)、产黄青霉、草酸青霉(P. oxalicum)、橘青霉(P. citrinum)、里氏木霉(Trichodermareesei)、深绿木霉(T. atroviride)、丛毛红曲霉(Monascuspilosus)、红色红曲霉(M. ruber)、灰葡萄孢菌(Botrytis cinerea)、藤仓镰孢菌(Gibberella fujikuroi)、轮枝镰刀菌(Fusarium verticillioide)、禾旋孢腔菌(Cochliobolus sativus)和玉米小斑病菌(Helminthosporium maydis)中被相继报道[2].研究发现laeA能够正调控包括抗生素和真菌毒素在内的大量次级代谢物的生物合成,同时影响真菌的形态分化和发育过程[3].LaeA还能够通过刺激沉默基因的表达激活隐性代谢物的产生[4]. ...
Overexpression of the global regulator LaeA in Chaetomium globosum leads to the biosynthesis of Chaetoglobosin Z
2
2016
... id="C6">全局性调控是指由全局性调控因子参与并对多个次级代谢物生物合成以及形态分化进行的调控.在诱导丝状真菌产生次级代谢物时,外界环境刺激是一个很重要的因素,而全局性调控因子在其中起着关键作用.LaeA是近年来发现的一种丝状真菌全局性调控因子.2004年,美国威斯康星大学Keller研究组首次在构巢曲霉(Aspergillus nidulans)中分离到laeA基因,生物信息分析表明LaeA含有一个保守的参与组蛋白甲基化的S-腺苷甲硫氨酸(SAM)结合结构域[1],同时发现laeA的启动子区含有途径特异性调控蛋白AflR的结合位点.随后laeA在烟曲霉(A. funigatus)、黄曲霉(A. flavus)、米曲霉(A. oryzae)、黑曲霉(A. niger)、产黄青霉、草酸青霉(P. oxalicum)、橘青霉(P. citrinum)、里氏木霉(Trichodermareesei)、深绿木霉(T. atroviride)、丛毛红曲霉(Monascuspilosus)、红色红曲霉(M. ruber)、灰葡萄孢菌(Botrytis cinerea)、藤仓镰孢菌(Gibberella fujikuroi)、轮枝镰刀菌(Fusarium verticillioide)、禾旋孢腔菌(Cochliobolus sativus)和玉米小斑病菌(Helminthosporium maydis)中被相继报道[2].研究发现laeA能够正调控包括抗生素和真菌毒素在内的大量次级代谢物的生物合成,同时影响真菌的形态分化和发育过程[3].LaeA还能够通过刺激沉默基因的表达激活隐性代谢物的产生[4]. ...
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
Overexpression of LaeA enhances mevastatin production and reduces sporulation of Penicillium citrinum
1
2014
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
橘青霉全局性调控因子LaeA相互作用蛋白研究 [学位论文]
1
2014
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
橘青霉全局性调控因子LaeA相互作用蛋白研究 [学位论文]
1
2014
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
Putative methyltransferase LaeA and transcription factor CreA are necessary for proper asexual development and controlling secondary metabolic gene cluster expression
1
2016
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
Inactivation of the global regulator LaeA in Monascus ruber results in a species-dependent response in sporulation and secondary metabolism
1
2016
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
黑曲霉FGSC A1279次级代谢调控研究[学位论文]
1
2014
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
黑曲霉FGSC A1279次级代谢调控研究[学位论文]
1
2014
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
Profiling of secondary metabolite gene clusters regulated by LaeA in Aspergillus niger FGSC A1279 based on genome sequencing and transcriptome analysis
1
2018
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
Distinct roles of velvet complex in the development, stress tolerance, and secondary metabolism in Pestalotiopsis microspora, a taxol producer
3
2018
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
... [11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
Secondary metabolism: regulation and role in fungal biology
1
2008
... id="C7">西南大学胡昌华课题组发现laeA能够通过促进橘青霉中途径特异性调控基因mlcR和结构基因mlcB的转录正调控美伐他汀的合成,因此可以通过高表达laeA来提高美伐他汀的产量[5].他们还发现laeA负调控橘青霉孢子的形成,并鉴定了3种与LaeA互作的蛋白.其中Sey-1可能参与调控G蛋白信号通路,DST1-like蛋白可能参与次级代谢的调控,胱硫醚-β-合成酶可能与LaeA组蛋白的甲基修饰相关[6].山东大学曲音波课题组发现laeA在发酵前期负调控草酸青霉中纤维素酶的合成,在中后期则正调控纤维素酶的合成,而且是发酵后期其他转录调控因子调控纤维素酶基因表达所必需的[7].华中农业大学陈福生课题组发现laeA除了调控红曲霉的菌落直径、孢子萌发和分生孢子形成外,还能正调控包括色素和橘霉素在内的6种已知次级代谢物的生物合成[8].华南理工大学潘力课题组通过转录组分析,发现laeA在转录水平正调控黑曲霉次级代谢生物合成基因簇中大约27%的基因表达,证明laeA普遍影响丝状真菌次级代谢[9, 10].北京师范大学朱旭东课题组发现laeA正调控球毛壳中pestalotiollide B的合成[11].中国医学科学院药用植物研究所邹忠梅课题组发现过表达laeA能够激活球毛壳菌新化合物Chaetoglobosin Z的产生[4],为隐性次级代谢物的激活提供了新的策略.然而,laeA调控次级代谢的作用机制至今尚不清楚.目前,推测有以下3种可能的作用机制:(1) LaeA作为甲基转移酶修饰组蛋白和其他蛋白的甲基化水平,通过影响染色体的状态而改变次级代谢物的合成[12];(2) LaeA通过与途径特异性调控因子作用从而共同调控次级代谢物的合成[1];(3) LaeA通过G蛋白信号转导途径将次级代谢和形态分化联系起来[1]. ...
The velvet family of fungal regulators contains a DNA- binding domain structurally similar to NF-κB
1
2013
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
The VeA regulatory system and its role in morphological and chemical development in fungi
1
2008
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
The veA gene activates sexual development in Aspergillus nidulans
1
2002
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
The expression of sterigmatocystin and penicillin genes in Aspergillus nidulans is controlled by veA, a gene required for sexual development
1
2003
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism
1
2008
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
橘青霉中全局性调控基因Pci-veA的克隆鉴定及其特性的初步研究[学位论文]
1
2011
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
橘青霉中全局性调控基因Pci-veA的克隆鉴定及其特性的初步研究[学位论文]
1
2011
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
橘青霉veA的全局性调控作用及其分子机制 [学位论文]
1
2015
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
橘青霉veA的全局性调控作用及其分子机制 [学位论文]
1
2015
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
Involvement of a velvet protein FgVeA in the regulation of asexual development, lipid and secondary metabolisms and virulence in Fusarium graminearum
1
2011
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
FgVELB is associated with vegetative differentiation, secondary metabolism and virulence in Fusarium graminearum
1
2012
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
Coordinated and distinct functions of velvet proteins in Fusarium verticillioides
1
2014
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
The regulatory gene VosA affects conidiogenesis and is involved in virulence of the fungal cereal pathogen Cochliobolus sativus
1
2015
... id="C8">Velvet蛋白家族是一类真菌特有的、含有Velvet 结构域的蛋白,包括VeA、VelB、VosA和VelC.其中VeA是在构巢曲霉中首次发现的Velvet类光敏蛋白,其N端含有一个保守的核定位信号,C端含有一个PEST区域[13].Velvet蛋白家族在丝状真菌中的主要功能是调控形态分化和次级代谢, 并且受到光、温度等环境因素影响[14].VeA除了影响丝状真菌的形态分化和次级代谢外,还正调控有性生殖和负调控无性生殖[15].此外,VeA还能激活隐性基因簇[16].VeA通过与VelB和LaeA形成蛋白复合体,共同调控丝状真菌形态分化和次级代谢[17].我国学者也在此领域开展了相关的研究,发现橘青霉中VeA能够抑制菌体的无性发育,并通过促进美伐他汀簇内结构基因和调控基因的转录提高美伐他汀的产量;VeA能够增强菌体对离子和氧化胁迫的耐受性[18, 19].与大多数VeA抑制无性生殖和激活次级代谢物合成不同,球毛壳中VeA正调控分生孢子的产生和菌丝的生长,但负调控pestalotiollide B的合成,这暗示着球毛壳中VeA对次级代谢和形态分化的作用机制具有特殊性[11].浙江大学马忠华课题组发现禾谷镰刀菌中FgVeA正调控脱氧雪腐镰刀菌烯醇(DON)的产生,并参与细胞内脂滴的合成.他们通过酵母双杂交获得了6个与FgVeA互作的蛋白,分析发现均含有甲基转移酶结构域.而这些Velvet复合体蛋白与构巢曲霉中相对应的蛋白作用完全不同,暗示着禾谷镰刀菌中Velvet蛋白复合体可能具有新的作用机制[20].FgVelB的破坏增加了禾谷镰刀菌对渗透压的抗性,阻碍了气生菌丝的生长,减弱了菌丝的疏水性,增加了分生孢子量;另外FgVelB还正调控DON的产生,最终导致FgVelB突变株对开花麦穗致病性的减弱[21].球毛壳菌VelB在正调控pestalotiollide B合成的同时,也调控了菌体的形态分化以及菌体对渗透压的敏感性[11].中国科学院微生物研究所李少杰课题组发现,轮枝镰刀菌中Velvet相关蛋白FvVE1、FvVelB和FvVelC能够形成复合物而发挥功能,FvVE1或FvVelB的敲除阻碍了小分生孢子的产生,但激活了大分生孢子的形成,同时发现二者均正调控烟曲霉毒素的产生.FvVE1和FvVelB参与调控菌体对氧化胁迫的耐受性,但FvVelC并不参与形态分化和次级代谢这两个重要的生物学过 程[22].而我国学者对VosA和VelC的研究较少,仅发现VosA参与禾旋孢腔菌分生孢子产生和毒力过程[23],目前尚未见关于其参与次级代谢的报道. ...
Roles of phospholipid methyltransferases in pycnidia development, stress tolerance and secondary metabolism in the taxol-producing fungus Pestalotiopsis microspore
1
2018
... id="C9">除了LaeA和Velvet之外,其他全局性调控因子与次级代谢之间的关系也有报道.例如,朱旭东课题组发现小孢拟盘多毛孢(Pestalotiopsis microspora)中磷脂甲基转移酶基因choA和choC的缺失使得菌体呈现出异常的形态分化,而choA和choC在次级代谢中的作用则完全不同[24].华东理工大学张元兴课题组发现海洋来源的灰绿曲霉调控蛋白Agseb1与酿酒酵母中的调控蛋白Msn2/4具有较高的同源性,Agseb1除了能够应对胁迫和调节菌体的形态分化之外,Agseb1缺失株中aspergiolide A的产量降低了43%[25].西南大学范艳华课题组发现球孢白僵菌(Beauveria bassiana)中产生的卵孢素通过抑制死亡虫体上细菌的大量增殖来保障真菌获得营养物质,最终完成菌丝生长和产孢过程.而球孢白僵菌中全局性调控因子BbSmr1通过负调控生物合成基因簇中转录因子OpS3的表达负调控卵孢素的合成[26].该项工作第一次向人们阐述了卵孢素如何帮助白僵菌在与细菌争夺死亡虫体营养的过程中获得优势.淡紫拟青霉(Paecilomyces lilacinus)产生的具有杀线虫活性的白灰制菌素在生物防治上具有重要的应用价值.华中农业大学肖炎农课题组发现淡紫拟青霉中RolP的缺失导致白灰制菌素不再产生,高表达rolP则可以提高白灰制菌素的产量,同时发现rolP与对线虫的毒性呈正相关[27].本课题组在研究头孢菌素生物合成的调控过程中,发现影响菌丝分隔的AcsepH基因和发育调控基因AcstuA的缺失都会导致顶头孢霉(Acremonium chrysogenum)抗生素产量急剧下降[28, 29],而一个Myb家族转录因子编码基因AcmybA的缺失则显著增加了分生孢子和头孢菌素的产生[30],说明次级代谢与形态分化的紧密联系在丝状真菌中是普遍存在的. ...
Transcription factor Agseb1 affects development, osmotic stress response, and secondary metabolism in marine- derived Aspergillus glaucus
1
2017
... id="C9">除了LaeA和Velvet之外,其他全局性调控因子与次级代谢之间的关系也有报道.例如,朱旭东课题组发现小孢拟盘多毛孢(Pestalotiopsis microspora)中磷脂甲基转移酶基因choA和choC的缺失使得菌体呈现出异常的形态分化,而choA和choC在次级代谢中的作用则完全不同[24].华东理工大学张元兴课题组发现海洋来源的灰绿曲霉调控蛋白Agseb1与酿酒酵母中的调控蛋白Msn2/4具有较高的同源性,Agseb1除了能够应对胁迫和调节菌体的形态分化之外,Agseb1缺失株中aspergiolide A的产量降低了43%[25].西南大学范艳华课题组发现球孢白僵菌(Beauveria bassiana)中产生的卵孢素通过抑制死亡虫体上细菌的大量增殖来保障真菌获得营养物质,最终完成菌丝生长和产孢过程.而球孢白僵菌中全局性调控因子BbSmr1通过负调控生物合成基因簇中转录因子OpS3的表达负调控卵孢素的合成[26].该项工作第一次向人们阐述了卵孢素如何帮助白僵菌在与细菌争夺死亡虫体营养的过程中获得优势.淡紫拟青霉(Paecilomyces lilacinus)产生的具有杀线虫活性的白灰制菌素在生物防治上具有重要的应用价值.华中农业大学肖炎农课题组发现淡紫拟青霉中RolP的缺失导致白灰制菌素不再产生,高表达rolP则可以提高白灰制菌素的产量,同时发现rolP与对线虫的毒性呈正相关[27].本课题组在研究头孢菌素生物合成的调控过程中,发现影响菌丝分隔的AcsepH基因和发育调控基因AcstuA的缺失都会导致顶头孢霉(Acremonium chrysogenum)抗生素产量急剧下降[28, 29],而一个Myb家族转录因子编码基因AcmybA的缺失则显著增加了分生孢子和头孢菌素的产生[30],说明次级代谢与形态分化的紧密联系在丝状真菌中是普遍存在的. ...
Regulatory cascade and biological activity of Beauveria bassiana oosporein that limits bacterial growth after host death
2
2017
... id="C9">除了LaeA和Velvet之外,其他全局性调控因子与次级代谢之间的关系也有报道.例如,朱旭东课题组发现小孢拟盘多毛孢(Pestalotiopsis microspora)中磷脂甲基转移酶基因choA和choC的缺失使得菌体呈现出异常的形态分化,而choA和choC在次级代谢中的作用则完全不同[24].华东理工大学张元兴课题组发现海洋来源的灰绿曲霉调控蛋白Agseb1与酿酒酵母中的调控蛋白Msn2/4具有较高的同源性,Agseb1除了能够应对胁迫和调节菌体的形态分化之外,Agseb1缺失株中aspergiolide A的产量降低了43%[25].西南大学范艳华课题组发现球孢白僵菌(Beauveria bassiana)中产生的卵孢素通过抑制死亡虫体上细菌的大量增殖来保障真菌获得营养物质,最终完成菌丝生长和产孢过程.而球孢白僵菌中全局性调控因子BbSmr1通过负调控生物合成基因簇中转录因子OpS3的表达负调控卵孢素的合成[26].该项工作第一次向人们阐述了卵孢素如何帮助白僵菌在与细菌争夺死亡虫体营养的过程中获得优势.淡紫拟青霉(Paecilomyces lilacinus)产生的具有杀线虫活性的白灰制菌素在生物防治上具有重要的应用价值.华中农业大学肖炎农课题组发现淡紫拟青霉中RolP的缺失导致白灰制菌素不再产生,高表达rolP则可以提高白灰制菌素的产量,同时发现rolP与对线虫的毒性呈正相关[27].本课题组在研究头孢菌素生物合成的调控过程中,发现影响菌丝分隔的AcsepH基因和发育调控基因AcstuA的缺失都会导致顶头孢霉(Acremonium chrysogenum)抗生素产量急剧下降[28, 29],而一个Myb家族转录因子编码基因AcmybA的缺失则显著增加了分生孢子和头孢菌素的产生[30],说明次级代谢与形态分化的紧密联系在丝状真菌中是普遍存在的. ...
... id="C26">丝状真菌次级代谢具有多层次调控的特点,主要包括转录水平调控、表观遗传调控和翻译后水平调控等,最终形成一个具有高度组织性且十分精密的调控网络(图2).丝状真菌次级代谢分子调控领域,我国从无到有,目前已取得了长足的进步.基于丝状真菌与宿主相互作用的理念,范艳华课题组在国际上首次揭示了卵孢素通过抑制死亡虫体上的细菌来竞争营养物质[26].丝状真菌隐性基因簇激活和新型化合物挖掘是国际上新兴的研究领域,通过异源表达、定向遗传改造、表观遗传修饰以及物种间互作等,能够获得多种新结构代谢产物(图3).我国科研人员也积极开展了相关工作,毛旭明和胡友财课题组通过表观遗传的改变快速同时激活齿梗孢霉多个隐性基因簇[38],尹文兵课题组先后在无花果拟盘多毛孢和罗氏绿僵菌通过隐性基因簇激活获得了多种新结构化合物[42, 43].越来越多的研究成果极大地促进了我国在丝状真菌次级代谢调控领域的国际竞争力. ...
II)2Cys6 putative transcription factor is involved in the regulation of leucinostatin production and pathogenicity of the nematophagous fungus Paecilomyces lilacinus
1
2015
... id="C9">除了LaeA和Velvet之外,其他全局性调控因子与次级代谢之间的关系也有报道.例如,朱旭东课题组发现小孢拟盘多毛孢(Pestalotiopsis microspora)中磷脂甲基转移酶基因choA和choC的缺失使得菌体呈现出异常的形态分化,而choA和choC在次级代谢中的作用则完全不同[24].华东理工大学张元兴课题组发现海洋来源的灰绿曲霉调控蛋白Agseb1与酿酒酵母中的调控蛋白Msn2/4具有较高的同源性,Agseb1除了能够应对胁迫和调节菌体的形态分化之外,Agseb1缺失株中aspergiolide A的产量降低了43%[25].西南大学范艳华课题组发现球孢白僵菌(Beauveria bassiana)中产生的卵孢素通过抑制死亡虫体上细菌的大量增殖来保障真菌获得营养物质,最终完成菌丝生长和产孢过程.而球孢白僵菌中全局性调控因子BbSmr1通过负调控生物合成基因簇中转录因子OpS3的表达负调控卵孢素的合成[26].该项工作第一次向人们阐述了卵孢素如何帮助白僵菌在与细菌争夺死亡虫体营养的过程中获得优势.淡紫拟青霉(Paecilomyces lilacinus)产生的具有杀线虫活性的白灰制菌素在生物防治上具有重要的应用价值.华中农业大学肖炎农课题组发现淡紫拟青霉中RolP的缺失导致白灰制菌素不再产生,高表达rolP则可以提高白灰制菌素的产量,同时发现rolP与对线虫的毒性呈正相关[27].本课题组在研究头孢菌素生物合成的调控过程中,发现影响菌丝分隔的AcsepH基因和发育调控基因AcstuA的缺失都会导致顶头孢霉(Acremonium chrysogenum)抗生素产量急剧下降[28, 29],而一个Myb家族转录因子编码基因AcmybA的缺失则显著增加了分生孢子和头孢菌素的产生[30],说明次级代谢与形态分化的紧密联系在丝状真菌中是普遍存在的. ...
A septation related gene AcsepH in Acremonium chrysogenum is involved in the cellular differentiation and cephalosporin production
1
2013
... id="C9">除了LaeA和Velvet之外,其他全局性调控因子与次级代谢之间的关系也有报道.例如,朱旭东课题组发现小孢拟盘多毛孢(Pestalotiopsis microspora)中磷脂甲基转移酶基因choA和choC的缺失使得菌体呈现出异常的形态分化,而choA和choC在次级代谢中的作用则完全不同[24].华东理工大学张元兴课题组发现海洋来源的灰绿曲霉调控蛋白Agseb1与酿酒酵母中的调控蛋白Msn2/4具有较高的同源性,Agseb1除了能够应对胁迫和调节菌体的形态分化之外,Agseb1缺失株中aspergiolide A的产量降低了43%[25].西南大学范艳华课题组发现球孢白僵菌(Beauveria bassiana)中产生的卵孢素通过抑制死亡虫体上细菌的大量增殖来保障真菌获得营养物质,最终完成菌丝生长和产孢过程.而球孢白僵菌中全局性调控因子BbSmr1通过负调控生物合成基因簇中转录因子OpS3的表达负调控卵孢素的合成[26].该项工作第一次向人们阐述了卵孢素如何帮助白僵菌在与细菌争夺死亡虫体营养的过程中获得优势.淡紫拟青霉(Paecilomyces lilacinus)产生的具有杀线虫活性的白灰制菌素在生物防治上具有重要的应用价值.华中农业大学肖炎农课题组发现淡紫拟青霉中RolP的缺失导致白灰制菌素不再产生,高表达rolP则可以提高白灰制菌素的产量,同时发现rolP与对线虫的毒性呈正相关[27].本课题组在研究头孢菌素生物合成的调控过程中,发现影响菌丝分隔的AcsepH基因和发育调控基因AcstuA的缺失都会导致顶头孢霉(Acremonium chrysogenum)抗生素产量急剧下降[28, 29],而一个Myb家族转录因子编码基因AcmybA的缺失则显著增加了分生孢子和头孢菌素的产生[30],说明次级代谢与形态分化的紧密联系在丝状真菌中是普遍存在的. ...
AcstuA, which encodes an APSES transcription regulator, is involved in conidiation, cephalosporin biosynthesis and cell wall integrity of Acremonium chrysogenum
1
2015
... id="C9">除了LaeA和Velvet之外,其他全局性调控因子与次级代谢之间的关系也有报道.例如,朱旭东课题组发现小孢拟盘多毛孢(Pestalotiopsis microspora)中磷脂甲基转移酶基因choA和choC的缺失使得菌体呈现出异常的形态分化,而choA和choC在次级代谢中的作用则完全不同[24].华东理工大学张元兴课题组发现海洋来源的灰绿曲霉调控蛋白Agseb1与酿酒酵母中的调控蛋白Msn2/4具有较高的同源性,Agseb1除了能够应对胁迫和调节菌体的形态分化之外,Agseb1缺失株中aspergiolide A的产量降低了43%[25].西南大学范艳华课题组发现球孢白僵菌(Beauveria bassiana)中产生的卵孢素通过抑制死亡虫体上细菌的大量增殖来保障真菌获得营养物质,最终完成菌丝生长和产孢过程.而球孢白僵菌中全局性调控因子BbSmr1通过负调控生物合成基因簇中转录因子OpS3的表达负调控卵孢素的合成[26].该项工作第一次向人们阐述了卵孢素如何帮助白僵菌在与细菌争夺死亡虫体营养的过程中获得优势.淡紫拟青霉(Paecilomyces lilacinus)产生的具有杀线虫活性的白灰制菌素在生物防治上具有重要的应用价值.华中农业大学肖炎农课题组发现淡紫拟青霉中RolP的缺失导致白灰制菌素不再产生,高表达rolP则可以提高白灰制菌素的产量,同时发现rolP与对线虫的毒性呈正相关[27].本课题组在研究头孢菌素生物合成的调控过程中,发现影响菌丝分隔的AcsepH基因和发育调控基因AcstuA的缺失都会导致顶头孢霉(Acremonium chrysogenum)抗生素产量急剧下降[28, 29],而一个Myb家族转录因子编码基因AcmybA的缺失则显著增加了分生孢子和头孢菌素的产生[30],说明次级代谢与形态分化的紧密联系在丝状真菌中是普遍存在的. ...
A Myb transcription factor represses conidiation and cephalosporin C production in Acremonium chrysogenum
1
2018
... id="C9">除了LaeA和Velvet之外,其他全局性调控因子与次级代谢之间的关系也有报道.例如,朱旭东课题组发现小孢拟盘多毛孢(Pestalotiopsis microspora)中磷脂甲基转移酶基因choA和choC的缺失使得菌体呈现出异常的形态分化,而choA和choC在次级代谢中的作用则完全不同[24].华东理工大学张元兴课题组发现海洋来源的灰绿曲霉调控蛋白Agseb1与酿酒酵母中的调控蛋白Msn2/4具有较高的同源性,Agseb1除了能够应对胁迫和调节菌体的形态分化之外,Agseb1缺失株中aspergiolide A的产量降低了43%[25].西南大学范艳华课题组发现球孢白僵菌(Beauveria bassiana)中产生的卵孢素通过抑制死亡虫体上细菌的大量增殖来保障真菌获得营养物质,最终完成菌丝生长和产孢过程.而球孢白僵菌中全局性调控因子BbSmr1通过负调控生物合成基因簇中转录因子OpS3的表达负调控卵孢素的合成[26].该项工作第一次向人们阐述了卵孢素如何帮助白僵菌在与细菌争夺死亡虫体营养的过程中获得优势.淡紫拟青霉(Paecilomyces lilacinus)产生的具有杀线虫活性的白灰制菌素在生物防治上具有重要的应用价值.华中农业大学肖炎农课题组发现淡紫拟青霉中RolP的缺失导致白灰制菌素不再产生,高表达rolP则可以提高白灰制菌素的产量,同时发现rolP与对线虫的毒性呈正相关[27].本课题组在研究头孢菌素生物合成的调控过程中,发现影响菌丝分隔的AcsepH基因和发育调控基因AcstuA的缺失都会导致顶头孢霉(Acremonium chrysogenum)抗生素产量急剧下降[28, 29],而一个Myb家族转录因子编码基因AcmybA的缺失则显著增加了分生孢子和头孢菌素的产生[30],说明次级代谢与形态分化的紧密联系在丝状真菌中是普遍存在的. ...
Regulation of fungal secondary metabolism
1
2013
... id="C10">丝状真菌次级代谢除了受到全局性调控之外,还受到途径特异性调控.途径特异性调控是指参与调控的基因位于次级代谢物生物合成基因簇内,其编码产物直接调控基因簇内基因的转录.研究发现,60%的真菌次级代谢物合成基因簇内含有途径特异性调控基因[31].在丝状真菌中发现的大多数途径特异性调控基因均正调控次级代谢物的生物合成.近年来的研究表明,途径特异性调控因子不仅可以调控所在基因簇中基因的转录,也可以调控基因簇外基因的转录[32],同时,不同的途径特异性调控因子之间可以进行交互调控. ...
Activation of a silent fungal polyketide biosynthesis pathway through regulatory cross talk with a cryptic nonribosomal peptide synthetase gene cluster
1
2010
... id="C10">丝状真菌次级代谢除了受到全局性调控之外,还受到途径特异性调控.途径特异性调控是指参与调控的基因位于次级代谢物生物合成基因簇内,其编码产物直接调控基因簇内基因的转录.研究发现,60%的真菌次级代谢物合成基因簇内含有途径特异性调控基因[31].在丝状真菌中发现的大多数途径特异性调控基因均正调控次级代谢物的生物合成.近年来的研究表明,途径特异性调控因子不仅可以调控所在基因簇中基因的转录,也可以调控基因簇外基因的转录[32],同时,不同的途径特异性调控因子之间可以进行交互调控. ...
Biosynthesis of antibiotic leucinostatins in bio-control fungus Purpureocillium lilacinum and their inhibition on phytophthora revealed by genome mining
1
2016
... id="C11">途径特异性调控相对简单和直接.中国农业科学院蔬菜花卉研究所谢丙炎课题组与中国科学院微生物研究所尹文兵课题组通过基因组分析、基因敲除等首次鉴定了白灰制菌素生物合成基因簇,并对其中途径特异性调控基因lcsF开展了研究,高表达lcsF使白灰制菌素产量增加了1.5倍[33].Verticillins是由罗杰斯无性穗霉(Clonostachys rogersoniana)产生的多硫代二酮哌嗪(ETP)类化合物,具有较强的细胞毒活性和成药前景.本课题组与中国人民解放军军事医学科学院车永胜课题组合作,克隆并通过实验确证了verticillin生物合成基因簇[34].该基因簇中存在一个途径特异性调控基因verZ,其编码产物通过直接与结构基因启动子区结合正调控verticillin的生物合成[35].Verticillin生物合成基因簇的克隆及其调控机制的阐明,为该化合物的定向改造及利用奠定了理论基础.西北农林科技大学许金荣课题组发现禾谷镰刀菌途径特异性调控基因Tri6和Tri10正调控DON的合成,而加入外源cAMP能够部分回补Tri10突变株中DON的产生,但对Tri6突变引起的DON缺失没有回补作用,暗示Tri6在参与cAMP- PKA途径介导的DON合成过程中起到至关重要的作用[36],这也是首次证明外源cAMP通过Tri10调控真菌毒素DON的合成. ...
Identification and characterization of the verticillin biosynthetic gene cluster in Clonostachys rogersoniana
1
2017
... id="C11">途径特异性调控相对简单和直接.中国农业科学院蔬菜花卉研究所谢丙炎课题组与中国科学院微生物研究所尹文兵课题组通过基因组分析、基因敲除等首次鉴定了白灰制菌素生物合成基因簇,并对其中途径特异性调控基因lcsF开展了研究,高表达lcsF使白灰制菌素产量增加了1.5倍[33].Verticillins是由罗杰斯无性穗霉(Clonostachys rogersoniana)产生的多硫代二酮哌嗪(ETP)类化合物,具有较强的细胞毒活性和成药前景.本课题组与中国人民解放军军事医学科学院车永胜课题组合作,克隆并通过实验确证了verticillin生物合成基因簇[34].该基因簇中存在一个途径特异性调控基因verZ,其编码产物通过直接与结构基因启动子区结合正调控verticillin的生物合成[35].Verticillin生物合成基因簇的克隆及其调控机制的阐明,为该化合物的定向改造及利用奠定了理论基础.西北农林科技大学许金荣课题组发现禾谷镰刀菌途径特异性调控基因Tri6和Tri10正调控DON的合成,而加入外源cAMP能够部分回补Tri10突变株中DON的产生,但对Tri6突变引起的DON缺失没有回补作用,暗示Tri6在参与cAMP- PKA途径介导的DON合成过程中起到至关重要的作用[36],这也是首次证明外源cAMP通过Tri10调控真菌毒素DON的合成. ...
VerZ, a Zn(II)2Cys6 DNA-binding protein, regulates the biosynthesis of verticillin in Clonostachys rogersoniana
1
2017
... id="C11">途径特异性调控相对简单和直接.中国农业科学院蔬菜花卉研究所谢丙炎课题组与中国科学院微生物研究所尹文兵课题组通过基因组分析、基因敲除等首次鉴定了白灰制菌素生物合成基因簇,并对其中途径特异性调控基因lcsF开展了研究,高表达lcsF使白灰制菌素产量增加了1.5倍[33].Verticillins是由罗杰斯无性穗霉(Clonostachys rogersoniana)产生的多硫代二酮哌嗪(ETP)类化合物,具有较强的细胞毒活性和成药前景.本课题组与中国人民解放军军事医学科学院车永胜课题组合作,克隆并通过实验确证了verticillin生物合成基因簇[34].该基因簇中存在一个途径特异性调控基因verZ,其编码产物通过直接与结构基因启动子区结合正调控verticillin的生物合成[35].Verticillin生物合成基因簇的克隆及其调控机制的阐明,为该化合物的定向改造及利用奠定了理论基础.西北农林科技大学许金荣课题组发现禾谷镰刀菌途径特异性调控基因Tri6和Tri10正调控DON的合成,而加入外源cAMP能够部分回补Tri10突变株中DON的产生,但对Tri6突变引起的DON缺失没有回补作用,暗示Tri6在参与cAMP- PKA途径介导的DON合成过程中起到至关重要的作用[36],这也是首次证明外源cAMP通过Tri10调控真菌毒素DON的合成. ...
TRI6 and TRI10 play different roles in the regulation of deoxynivalenol (DON) production by cAMPsignalling in Fusarium graminearum
1
2016
... id="C11">途径特异性调控相对简单和直接.中国农业科学院蔬菜花卉研究所谢丙炎课题组与中国科学院微生物研究所尹文兵课题组通过基因组分析、基因敲除等首次鉴定了白灰制菌素生物合成基因簇,并对其中途径特异性调控基因lcsF开展了研究,高表达lcsF使白灰制菌素产量增加了1.5倍[33].Verticillins是由罗杰斯无性穗霉(Clonostachys rogersoniana)产生的多硫代二酮哌嗪(ETP)类化合物,具有较强的细胞毒活性和成药前景.本课题组与中国人民解放军军事医学科学院车永胜课题组合作,克隆并通过实验确证了verticillin生物合成基因簇[34].该基因簇中存在一个途径特异性调控基因verZ,其编码产物通过直接与结构基因启动子区结合正调控verticillin的生物合成[35].Verticillin生物合成基因簇的克隆及其调控机制的阐明,为该化合物的定向改造及利用奠定了理论基础.西北农林科技大学许金荣课题组发现禾谷镰刀菌途径特异性调控基因Tri6和Tri10正调控DON的合成,而加入外源cAMP能够部分回补Tri10突变株中DON的产生,但对Tri6突变引起的DON缺失没有回补作用,暗示Tri6在参与cAMP- PKA途径介导的DON合成过程中起到至关重要的作用[36],这也是首次证明外源cAMP通过Tri10调控真菌毒素DON的合成. ...
SchroeckhV. Fungal secondary metabolites- strategies to activate silent gene clusters
1
2011
... id="C12">丝状真菌次级代谢与其基因簇所处的表观遗传状态有着密切的关系.通过表观遗传操作能大范围改变代谢物谱,以及激活隐性次级代谢物的产生[37].表观遗传调控是一种基于核酸修饰的调控方式,主要包括DNA甲基化、组蛋白甲基化、乙酰化、磷酸化和泛素化等.其中,组蛋白甲基化是最稳定的修饰方式,因此最适合作为稳定的表观遗传信息.而乙酰化修饰则具有较高的动态性,同时磷酸化和泛素化修饰相对不太稳定,因此它们更为灵活地影响染色质的结构和功能.组蛋白遗传修饰之间相互关联并共同控制着基因的表达,从而最终影响丝状真菌次级代谢物的合成.其中,组蛋白甲基化和乙酰化研究的最为透彻.LaeA作为组蛋白甲基转移酶,通过调控组蛋白和改变染色体结构来调控次级代谢产物的合成.通常组蛋白甲基转移酶和组蛋白去乙酰化酶(HDAC)能够阻遏次级代谢物的合成.浙江大学毛旭明、中国医学科学院药物研究所胡友财课题组以及美国加州大学唐奕课题组合作研究发现,敲除齿梗孢霉(Calcarisporium arbuscula)中组蛋白去乙酰化酶使得菌体中超过75%的次级代谢基因簇表达水平上升,从该突变株中分离的10种化合物中4种具有新结构[38],该工作提示可以通过表观遗传的改变快速同时激活多个隐性基因簇,为基因组挖掘提供了强大的工具.朱旭东课题组发现小孢拟盘多毛孢组蛋白乙酰转移酶基因hat1正调控次级代谢和分生孢子的形成,同时也控制细胞壁的完整性[39].中国农业科学院植物保护研究所冯洁课题组发现敲除组蛋白乙酰转移酶基因FgSAS3和FgGCN5增加了禾谷镰刀菌对渗透压和活性氧(ROS)的敏感性,FgSAS3敲除株中分生孢子的量和子囊壳的形成减少,而FgGCN5敲除株则完全不能形成分生孢子和子囊壳.同时发现FgSAS3和FgGCN5是禾谷镰刀菌感染小麦穗和番茄果实的重要致病因子.FgSAS3和FgGCN敲除株不再产生DON,这一结果与敲除株中参与DON合成的关键基因的转录明显下降相一致[40].福建农林大学汪世华课题组发现黄曲霉中组蛋白乙酰转移酶基因AflgcnE的缺失抑制了黄曲霉的生长,降低了细胞表面的疏水性,同时使得菌体不能产生无性孢子和菌核,并导致黄曲霉毒素合成相关基因表达量急剧下降,黄曲霉毒素不再产生[41].尹文兵课题组发现敲除无花果拟盘多毛孢(Pestalotiopsis fici)中组蛋白甲基转移酶基因PfcclA和组蛋白去乙酰化酶基因PfhdaA激活了15种新的聚酮化合物的产生,同时提高了11种大环内酯类抗生素的产量[42].另外罗氏绿僵菌(Metarhizium robertsii)中hat1的敲除激活了孤儿次级代谢物生物合成基因的表达,在该敲除株中分离得到了11种新化合物[43].近年来的研究发现,细菌和真菌中还普遍存在组蛋白赖氨酸的琥珀酰化修饰,并且已经证明其在细胞分化、中心代谢等重要生命活动中发挥着重要作用,尤其是西南大学谢建平课题组对结核分枝杆菌琥珀酰化与其致病性关系进行了详细的蛋白组学研究,为结核病的诊断和防治提供了科学依据[44].然而,截至目前为止还没有在丝状真菌中发现琥珀酰化与次级代谢产物合成之间关系的报道.总之,上述研究证明表观遗传调控是发掘新化合物便捷和有效的策略之一. ...
Epigenetic genome mining of an endophytic fungus leads to the pleiotropic biosynthesis of natural products
2
2015
... id="C12">丝状真菌次级代谢与其基因簇所处的表观遗传状态有着密切的关系.通过表观遗传操作能大范围改变代谢物谱,以及激活隐性次级代谢物的产生[37].表观遗传调控是一种基于核酸修饰的调控方式,主要包括DNA甲基化、组蛋白甲基化、乙酰化、磷酸化和泛素化等.其中,组蛋白甲基化是最稳定的修饰方式,因此最适合作为稳定的表观遗传信息.而乙酰化修饰则具有较高的动态性,同时磷酸化和泛素化修饰相对不太稳定,因此它们更为灵活地影响染色质的结构和功能.组蛋白遗传修饰之间相互关联并共同控制着基因的表达,从而最终影响丝状真菌次级代谢物的合成.其中,组蛋白甲基化和乙酰化研究的最为透彻.LaeA作为组蛋白甲基转移酶,通过调控组蛋白和改变染色体结构来调控次级代谢产物的合成.通常组蛋白甲基转移酶和组蛋白去乙酰化酶(HDAC)能够阻遏次级代谢物的合成.浙江大学毛旭明、中国医学科学院药物研究所胡友财课题组以及美国加州大学唐奕课题组合作研究发现,敲除齿梗孢霉(Calcarisporium arbuscula)中组蛋白去乙酰化酶使得菌体中超过75%的次级代谢基因簇表达水平上升,从该突变株中分离的10种化合物中4种具有新结构[38],该工作提示可以通过表观遗传的改变快速同时激活多个隐性基因簇,为基因组挖掘提供了强大的工具.朱旭东课题组发现小孢拟盘多毛孢组蛋白乙酰转移酶基因hat1正调控次级代谢和分生孢子的形成,同时也控制细胞壁的完整性[39].中国农业科学院植物保护研究所冯洁课题组发现敲除组蛋白乙酰转移酶基因FgSAS3和FgGCN5增加了禾谷镰刀菌对渗透压和活性氧(ROS)的敏感性,FgSAS3敲除株中分生孢子的量和子囊壳的形成减少,而FgGCN5敲除株则完全不能形成分生孢子和子囊壳.同时发现FgSAS3和FgGCN5是禾谷镰刀菌感染小麦穗和番茄果实的重要致病因子.FgSAS3和FgGCN敲除株不再产生DON,这一结果与敲除株中参与DON合成的关键基因的转录明显下降相一致[40].福建农林大学汪世华课题组发现黄曲霉中组蛋白乙酰转移酶基因AflgcnE的缺失抑制了黄曲霉的生长,降低了细胞表面的疏水性,同时使得菌体不能产生无性孢子和菌核,并导致黄曲霉毒素合成相关基因表达量急剧下降,黄曲霉毒素不再产生[41].尹文兵课题组发现敲除无花果拟盘多毛孢(Pestalotiopsis fici)中组蛋白甲基转移酶基因PfcclA和组蛋白去乙酰化酶基因PfhdaA激活了15种新的聚酮化合物的产生,同时提高了11种大环内酯类抗生素的产量[42].另外罗氏绿僵菌(Metarhizium robertsii)中hat1的敲除激活了孤儿次级代谢物生物合成基因的表达,在该敲除株中分离得到了11种新化合物[43].近年来的研究发现,细菌和真菌中还普遍存在组蛋白赖氨酸的琥珀酰化修饰,并且已经证明其在细胞分化、中心代谢等重要生命活动中发挥着重要作用,尤其是西南大学谢建平课题组对结核分枝杆菌琥珀酰化与其致病性关系进行了详细的蛋白组学研究,为结核病的诊断和防治提供了科学依据[44].然而,截至目前为止还没有在丝状真菌中发现琥珀酰化与次级代谢产物合成之间关系的报道.总之,上述研究证明表观遗传调控是发掘新化合物便捷和有效的策略之一. ...
... id="C26">丝状真菌次级代谢具有多层次调控的特点,主要包括转录水平调控、表观遗传调控和翻译后水平调控等,最终形成一个具有高度组织性且十分精密的调控网络(图2).丝状真菌次级代谢分子调控领域,我国从无到有,目前已取得了长足的进步.基于丝状真菌与宿主相互作用的理念,范艳华课题组在国际上首次揭示了卵孢素通过抑制死亡虫体上的细菌来竞争营养物质[26].丝状真菌隐性基因簇激活和新型化合物挖掘是国际上新兴的研究领域,通过异源表达、定向遗传改造、表观遗传修饰以及物种间互作等,能够获得多种新结构代谢产物(图3).我国科研人员也积极开展了相关工作,毛旭明和胡友财课题组通过表观遗传的改变快速同时激活齿梗孢霉多个隐性基因簇[38],尹文兵课题组先后在无花果拟盘多毛孢和罗氏绿僵菌通过隐性基因簇激活获得了多种新结构化合物[42, 43].越来越多的研究成果极大地促进了我国在丝状真菌次级代谢调控领域的国际竞争力. ...
A B-type histone acetyltransferase Hat1 regulates secondary metabolism, conidiation, and cell wall integrity in the taxol-producing fungus Pestalotiopsis microspora
1
2016
... id="C12">丝状真菌次级代谢与其基因簇所处的表观遗传状态有着密切的关系.通过表观遗传操作能大范围改变代谢物谱,以及激活隐性次级代谢物的产生[37].表观遗传调控是一种基于核酸修饰的调控方式,主要包括DNA甲基化、组蛋白甲基化、乙酰化、磷酸化和泛素化等.其中,组蛋白甲基化是最稳定的修饰方式,因此最适合作为稳定的表观遗传信息.而乙酰化修饰则具有较高的动态性,同时磷酸化和泛素化修饰相对不太稳定,因此它们更为灵活地影响染色质的结构和功能.组蛋白遗传修饰之间相互关联并共同控制着基因的表达,从而最终影响丝状真菌次级代谢物的合成.其中,组蛋白甲基化和乙酰化研究的最为透彻.LaeA作为组蛋白甲基转移酶,通过调控组蛋白和改变染色体结构来调控次级代谢产物的合成.通常组蛋白甲基转移酶和组蛋白去乙酰化酶(HDAC)能够阻遏次级代谢物的合成.浙江大学毛旭明、中国医学科学院药物研究所胡友财课题组以及美国加州大学唐奕课题组合作研究发现,敲除齿梗孢霉(Calcarisporium arbuscula)中组蛋白去乙酰化酶使得菌体中超过75%的次级代谢基因簇表达水平上升,从该突变株中分离的10种化合物中4种具有新结构[38],该工作提示可以通过表观遗传的改变快速同时激活多个隐性基因簇,为基因组挖掘提供了强大的工具.朱旭东课题组发现小孢拟盘多毛孢组蛋白乙酰转移酶基因hat1正调控次级代谢和分生孢子的形成,同时也控制细胞壁的完整性[39].中国农业科学院植物保护研究所冯洁课题组发现敲除组蛋白乙酰转移酶基因FgSAS3和FgGCN5增加了禾谷镰刀菌对渗透压和活性氧(ROS)的敏感性,FgSAS3敲除株中分生孢子的量和子囊壳的形成减少,而FgGCN5敲除株则完全不能形成分生孢子和子囊壳.同时发现FgSAS3和FgGCN5是禾谷镰刀菌感染小麦穗和番茄果实的重要致病因子.FgSAS3和FgGCN敲除株不再产生DON,这一结果与敲除株中参与DON合成的关键基因的转录明显下降相一致[40].福建农林大学汪世华课题组发现黄曲霉中组蛋白乙酰转移酶基因AflgcnE的缺失抑制了黄曲霉的生长,降低了细胞表面的疏水性,同时使得菌体不能产生无性孢子和菌核,并导致黄曲霉毒素合成相关基因表达量急剧下降,黄曲霉毒素不再产生[41].尹文兵课题组发现敲除无花果拟盘多毛孢(Pestalotiopsis fici)中组蛋白甲基转移酶基因PfcclA和组蛋白去乙酰化酶基因PfhdaA激活了15种新的聚酮化合物的产生,同时提高了11种大环内酯类抗生素的产量[42].另外罗氏绿僵菌(Metarhizium robertsii)中hat1的敲除激活了孤儿次级代谢物生物合成基因的表达,在该敲除株中分离得到了11种新化合物[43].近年来的研究发现,细菌和真菌中还普遍存在组蛋白赖氨酸的琥珀酰化修饰,并且已经证明其在细胞分化、中心代谢等重要生命活动中发挥着重要作用,尤其是西南大学谢建平课题组对结核分枝杆菌琥珀酰化与其致病性关系进行了详细的蛋白组学研究,为结核病的诊断和防治提供了科学依据[44].然而,截至目前为止还没有在丝状真菌中发现琥珀酰化与次级代谢产物合成之间关系的报道.总之,上述研究证明表观遗传调控是发掘新化合物便捷和有效的策略之一. ...
The Fusarium graminearum histone acetyltransferases are important for morphogenesis, DON biosynthesis, and pathogenicity
1
2018
... id="C12">丝状真菌次级代谢与其基因簇所处的表观遗传状态有着密切的关系.通过表观遗传操作能大范围改变代谢物谱,以及激活隐性次级代谢物的产生[37].表观遗传调控是一种基于核酸修饰的调控方式,主要包括DNA甲基化、组蛋白甲基化、乙酰化、磷酸化和泛素化等.其中,组蛋白甲基化是最稳定的修饰方式,因此最适合作为稳定的表观遗传信息.而乙酰化修饰则具有较高的动态性,同时磷酸化和泛素化修饰相对不太稳定,因此它们更为灵活地影响染色质的结构和功能.组蛋白遗传修饰之间相互关联并共同控制着基因的表达,从而最终影响丝状真菌次级代谢物的合成.其中,组蛋白甲基化和乙酰化研究的最为透彻.LaeA作为组蛋白甲基转移酶,通过调控组蛋白和改变染色体结构来调控次级代谢产物的合成.通常组蛋白甲基转移酶和组蛋白去乙酰化酶(HDAC)能够阻遏次级代谢物的合成.浙江大学毛旭明、中国医学科学院药物研究所胡友财课题组以及美国加州大学唐奕课题组合作研究发现,敲除齿梗孢霉(Calcarisporium arbuscula)中组蛋白去乙酰化酶使得菌体中超过75%的次级代谢基因簇表达水平上升,从该突变株中分离的10种化合物中4种具有新结构[38],该工作提示可以通过表观遗传的改变快速同时激活多个隐性基因簇,为基因组挖掘提供了强大的工具.朱旭东课题组发现小孢拟盘多毛孢组蛋白乙酰转移酶基因hat1正调控次级代谢和分生孢子的形成,同时也控制细胞壁的完整性[39].中国农业科学院植物保护研究所冯洁课题组发现敲除组蛋白乙酰转移酶基因FgSAS3和FgGCN5增加了禾谷镰刀菌对渗透压和活性氧(ROS)的敏感性,FgSAS3敲除株中分生孢子的量和子囊壳的形成减少,而FgGCN5敲除株则完全不能形成分生孢子和子囊壳.同时发现FgSAS3和FgGCN5是禾谷镰刀菌感染小麦穗和番茄果实的重要致病因子.FgSAS3和FgGCN敲除株不再产生DON,这一结果与敲除株中参与DON合成的关键基因的转录明显下降相一致[40].福建农林大学汪世华课题组发现黄曲霉中组蛋白乙酰转移酶基因AflgcnE的缺失抑制了黄曲霉的生长,降低了细胞表面的疏水性,同时使得菌体不能产生无性孢子和菌核,并导致黄曲霉毒素合成相关基因表达量急剧下降,黄曲霉毒素不再产生[41].尹文兵课题组发现敲除无花果拟盘多毛孢(Pestalotiopsis fici)中组蛋白甲基转移酶基因PfcclA和组蛋白去乙酰化酶基因PfhdaA激活了15种新的聚酮化合物的产生,同时提高了11种大环内酯类抗生素的产量[42].另外罗氏绿僵菌(Metarhizium robertsii)中hat1的敲除激活了孤儿次级代谢物生物合成基因的表达,在该敲除株中分离得到了11种新化合物[43].近年来的研究发现,细菌和真菌中还普遍存在组蛋白赖氨酸的琥珀酰化修饰,并且已经证明其在细胞分化、中心代谢等重要生命活动中发挥着重要作用,尤其是西南大学谢建平课题组对结核分枝杆菌琥珀酰化与其致病性关系进行了详细的蛋白组学研究,为结核病的诊断和防治提供了科学依据[44].然而,截至目前为止还没有在丝状真菌中发现琥珀酰化与次级代谢产物合成之间关系的报道.总之,上述研究证明表观遗传调控是发掘新化合物便捷和有效的策略之一. ...
The Aspergillus flavus histone acetyl transferase AflGcnE regulates morphogenesis, aflatoxin biosynthesis, and pathogenicity
1
2016
... id="C12">丝状真菌次级代谢与其基因簇所处的表观遗传状态有着密切的关系.通过表观遗传操作能大范围改变代谢物谱,以及激活隐性次级代谢物的产生[37].表观遗传调控是一种基于核酸修饰的调控方式,主要包括DNA甲基化、组蛋白甲基化、乙酰化、磷酸化和泛素化等.其中,组蛋白甲基化是最稳定的修饰方式,因此最适合作为稳定的表观遗传信息.而乙酰化修饰则具有较高的动态性,同时磷酸化和泛素化修饰相对不太稳定,因此它们更为灵活地影响染色质的结构和功能.组蛋白遗传修饰之间相互关联并共同控制着基因的表达,从而最终影响丝状真菌次级代谢物的合成.其中,组蛋白甲基化和乙酰化研究的最为透彻.LaeA作为组蛋白甲基转移酶,通过调控组蛋白和改变染色体结构来调控次级代谢产物的合成.通常组蛋白甲基转移酶和组蛋白去乙酰化酶(HDAC)能够阻遏次级代谢物的合成.浙江大学毛旭明、中国医学科学院药物研究所胡友财课题组以及美国加州大学唐奕课题组合作研究发现,敲除齿梗孢霉(Calcarisporium arbuscula)中组蛋白去乙酰化酶使得菌体中超过75%的次级代谢基因簇表达水平上升,从该突变株中分离的10种化合物中4种具有新结构[38],该工作提示可以通过表观遗传的改变快速同时激活多个隐性基因簇,为基因组挖掘提供了强大的工具.朱旭东课题组发现小孢拟盘多毛孢组蛋白乙酰转移酶基因hat1正调控次级代谢和分生孢子的形成,同时也控制细胞壁的完整性[39].中国农业科学院植物保护研究所冯洁课题组发现敲除组蛋白乙酰转移酶基因FgSAS3和FgGCN5增加了禾谷镰刀菌对渗透压和活性氧(ROS)的敏感性,FgSAS3敲除株中分生孢子的量和子囊壳的形成减少,而FgGCN5敲除株则完全不能形成分生孢子和子囊壳.同时发现FgSAS3和FgGCN5是禾谷镰刀菌感染小麦穗和番茄果实的重要致病因子.FgSAS3和FgGCN敲除株不再产生DON,这一结果与敲除株中参与DON合成的关键基因的转录明显下降相一致[40].福建农林大学汪世华课题组发现黄曲霉中组蛋白乙酰转移酶基因AflgcnE的缺失抑制了黄曲霉的生长,降低了细胞表面的疏水性,同时使得菌体不能产生无性孢子和菌核,并导致黄曲霉毒素合成相关基因表达量急剧下降,黄曲霉毒素不再产生[41].尹文兵课题组发现敲除无花果拟盘多毛孢(Pestalotiopsis fici)中组蛋白甲基转移酶基因PfcclA和组蛋白去乙酰化酶基因PfhdaA激活了15种新的聚酮化合物的产生,同时提高了11种大环内酯类抗生素的产量[42].另外罗氏绿僵菌(Metarhizium robertsii)中hat1的敲除激活了孤儿次级代谢物生物合成基因的表达,在该敲除株中分离得到了11种新化合物[43].近年来的研究发现,细菌和真菌中还普遍存在组蛋白赖氨酸的琥珀酰化修饰,并且已经证明其在细胞分化、中心代谢等重要生命活动中发挥着重要作用,尤其是西南大学谢建平课题组对结核分枝杆菌琥珀酰化与其致病性关系进行了详细的蛋白组学研究,为结核病的诊断和防治提供了科学依据[44].然而,截至目前为止还没有在丝状真菌中发现琥珀酰化与次级代谢产物合成之间关系的报道.总之,上述研究证明表观遗传调控是发掘新化合物便捷和有效的策略之一. ...
Polyketide production of pestaloficiols and macrodiolideficiolides revealed by manipulations of epigenetic regulators in an endophytic fungus
2
2016
... id="C12">丝状真菌次级代谢与其基因簇所处的表观遗传状态有着密切的关系.通过表观遗传操作能大范围改变代谢物谱,以及激活隐性次级代谢物的产生[37].表观遗传调控是一种基于核酸修饰的调控方式,主要包括DNA甲基化、组蛋白甲基化、乙酰化、磷酸化和泛素化等.其中,组蛋白甲基化是最稳定的修饰方式,因此最适合作为稳定的表观遗传信息.而乙酰化修饰则具有较高的动态性,同时磷酸化和泛素化修饰相对不太稳定,因此它们更为灵活地影响染色质的结构和功能.组蛋白遗传修饰之间相互关联并共同控制着基因的表达,从而最终影响丝状真菌次级代谢物的合成.其中,组蛋白甲基化和乙酰化研究的最为透彻.LaeA作为组蛋白甲基转移酶,通过调控组蛋白和改变染色体结构来调控次级代谢产物的合成.通常组蛋白甲基转移酶和组蛋白去乙酰化酶(HDAC)能够阻遏次级代谢物的合成.浙江大学毛旭明、中国医学科学院药物研究所胡友财课题组以及美国加州大学唐奕课题组合作研究发现,敲除齿梗孢霉(Calcarisporium arbuscula)中组蛋白去乙酰化酶使得菌体中超过75%的次级代谢基因簇表达水平上升,从该突变株中分离的10种化合物中4种具有新结构[38],该工作提示可以通过表观遗传的改变快速同时激活多个隐性基因簇,为基因组挖掘提供了强大的工具.朱旭东课题组发现小孢拟盘多毛孢组蛋白乙酰转移酶基因hat1正调控次级代谢和分生孢子的形成,同时也控制细胞壁的完整性[39].中国农业科学院植物保护研究所冯洁课题组发现敲除组蛋白乙酰转移酶基因FgSAS3和FgGCN5增加了禾谷镰刀菌对渗透压和活性氧(ROS)的敏感性,FgSAS3敲除株中分生孢子的量和子囊壳的形成减少,而FgGCN5敲除株则完全不能形成分生孢子和子囊壳.同时发现FgSAS3和FgGCN5是禾谷镰刀菌感染小麦穗和番茄果实的重要致病因子.FgSAS3和FgGCN敲除株不再产生DON,这一结果与敲除株中参与DON合成的关键基因的转录明显下降相一致[40].福建农林大学汪世华课题组发现黄曲霉中组蛋白乙酰转移酶基因AflgcnE的缺失抑制了黄曲霉的生长,降低了细胞表面的疏水性,同时使得菌体不能产生无性孢子和菌核,并导致黄曲霉毒素合成相关基因表达量急剧下降,黄曲霉毒素不再产生[41].尹文兵课题组发现敲除无花果拟盘多毛孢(Pestalotiopsis fici)中组蛋白甲基转移酶基因PfcclA和组蛋白去乙酰化酶基因PfhdaA激活了15种新的聚酮化合物的产生,同时提高了11种大环内酯类抗生素的产量[42].另外罗氏绿僵菌(Metarhizium robertsii)中hat1的敲除激活了孤儿次级代谢物生物合成基因的表达,在该敲除株中分离得到了11种新化合物[43].近年来的研究发现,细菌和真菌中还普遍存在组蛋白赖氨酸的琥珀酰化修饰,并且已经证明其在细胞分化、中心代谢等重要生命活动中发挥着重要作用,尤其是西南大学谢建平课题组对结核分枝杆菌琥珀酰化与其致病性关系进行了详细的蛋白组学研究,为结核病的诊断和防治提供了科学依据[44].然而,截至目前为止还没有在丝状真菌中发现琥珀酰化与次级代谢产物合成之间关系的报道.总之,上述研究证明表观遗传调控是发掘新化合物便捷和有效的策略之一. ...
... id="C26">丝状真菌次级代谢具有多层次调控的特点,主要包括转录水平调控、表观遗传调控和翻译后水平调控等,最终形成一个具有高度组织性且十分精密的调控网络(图2).丝状真菌次级代谢分子调控领域,我国从无到有,目前已取得了长足的进步.基于丝状真菌与宿主相互作用的理念,范艳华课题组在国际上首次揭示了卵孢素通过抑制死亡虫体上的细菌来竞争营养物质[26].丝状真菌隐性基因簇激活和新型化合物挖掘是国际上新兴的研究领域,通过异源表达、定向遗传改造、表观遗传修饰以及物种间互作等,能够获得多种新结构代谢产物(图3).我国科研人员也积极开展了相关工作,毛旭明和胡友财课题组通过表观遗传的改变快速同时激活齿梗孢霉多个隐性基因簇[38],尹文兵课题组先后在无花果拟盘多毛孢和罗氏绿僵菌通过隐性基因簇激活获得了多种新结构化合物[42, 43].越来越多的研究成果极大地促进了我国在丝状真菌次级代谢调控领域的国际竞争力. ...
Deletion of a histone acetyltransferase leads to the pleiotropic activation of natural products in Metarhizium robertsii
2
2017
... id="C12">丝状真菌次级代谢与其基因簇所处的表观遗传状态有着密切的关系.通过表观遗传操作能大范围改变代谢物谱,以及激活隐性次级代谢物的产生[37].表观遗传调控是一种基于核酸修饰的调控方式,主要包括DNA甲基化、组蛋白甲基化、乙酰化、磷酸化和泛素化等.其中,组蛋白甲基化是最稳定的修饰方式,因此最适合作为稳定的表观遗传信息.而乙酰化修饰则具有较高的动态性,同时磷酸化和泛素化修饰相对不太稳定,因此它们更为灵活地影响染色质的结构和功能.组蛋白遗传修饰之间相互关联并共同控制着基因的表达,从而最终影响丝状真菌次级代谢物的合成.其中,组蛋白甲基化和乙酰化研究的最为透彻.LaeA作为组蛋白甲基转移酶,通过调控组蛋白和改变染色体结构来调控次级代谢产物的合成.通常组蛋白甲基转移酶和组蛋白去乙酰化酶(HDAC)能够阻遏次级代谢物的合成.浙江大学毛旭明、中国医学科学院药物研究所胡友财课题组以及美国加州大学唐奕课题组合作研究发现,敲除齿梗孢霉(Calcarisporium arbuscula)中组蛋白去乙酰化酶使得菌体中超过75%的次级代谢基因簇表达水平上升,从该突变株中分离的10种化合物中4种具有新结构[38],该工作提示可以通过表观遗传的改变快速同时激活多个隐性基因簇,为基因组挖掘提供了强大的工具.朱旭东课题组发现小孢拟盘多毛孢组蛋白乙酰转移酶基因hat1正调控次级代谢和分生孢子的形成,同时也控制细胞壁的完整性[39].中国农业科学院植物保护研究所冯洁课题组发现敲除组蛋白乙酰转移酶基因FgSAS3和FgGCN5增加了禾谷镰刀菌对渗透压和活性氧(ROS)的敏感性,FgSAS3敲除株中分生孢子的量和子囊壳的形成减少,而FgGCN5敲除株则完全不能形成分生孢子和子囊壳.同时发现FgSAS3和FgGCN5是禾谷镰刀菌感染小麦穗和番茄果实的重要致病因子.FgSAS3和FgGCN敲除株不再产生DON,这一结果与敲除株中参与DON合成的关键基因的转录明显下降相一致[40].福建农林大学汪世华课题组发现黄曲霉中组蛋白乙酰转移酶基因AflgcnE的缺失抑制了黄曲霉的生长,降低了细胞表面的疏水性,同时使得菌体不能产生无性孢子和菌核,并导致黄曲霉毒素合成相关基因表达量急剧下降,黄曲霉毒素不再产生[41].尹文兵课题组发现敲除无花果拟盘多毛孢(Pestalotiopsis fici)中组蛋白甲基转移酶基因PfcclA和组蛋白去乙酰化酶基因PfhdaA激活了15种新的聚酮化合物的产生,同时提高了11种大环内酯类抗生素的产量[42].另外罗氏绿僵菌(Metarhizium robertsii)中hat1的敲除激活了孤儿次级代谢物生物合成基因的表达,在该敲除株中分离得到了11种新化合物[43].近年来的研究发现,细菌和真菌中还普遍存在组蛋白赖氨酸的琥珀酰化修饰,并且已经证明其在细胞分化、中心代谢等重要生命活动中发挥着重要作用,尤其是西南大学谢建平课题组对结核分枝杆菌琥珀酰化与其致病性关系进行了详细的蛋白组学研究,为结核病的诊断和防治提供了科学依据[44].然而,截至目前为止还没有在丝状真菌中发现琥珀酰化与次级代谢产物合成之间关系的报道.总之,上述研究证明表观遗传调控是发掘新化合物便捷和有效的策略之一. ...
... id="C26">丝状真菌次级代谢具有多层次调控的特点,主要包括转录水平调控、表观遗传调控和翻译后水平调控等,最终形成一个具有高度组织性且十分精密的调控网络(图2).丝状真菌次级代谢分子调控领域,我国从无到有,目前已取得了长足的进步.基于丝状真菌与宿主相互作用的理念,范艳华课题组在国际上首次揭示了卵孢素通过抑制死亡虫体上的细菌来竞争营养物质[26].丝状真菌隐性基因簇激活和新型化合物挖掘是国际上新兴的研究领域,通过异源表达、定向遗传改造、表观遗传修饰以及物种间互作等,能够获得多种新结构代谢产物(图3).我国科研人员也积极开展了相关工作,毛旭明和胡友财课题组通过表观遗传的改变快速同时激活齿梗孢霉多个隐性基因簇[38],尹文兵课题组先后在无花果拟盘多毛孢和罗氏绿僵菌通过隐性基因簇激活获得了多种新结构化合物[42, 43].越来越多的研究成果极大地促进了我国在丝状真菌次级代谢调控领域的国际竞争力. ...
结核分枝杆菌表观遗传与耐药新机理研究 [学位论文]
1
2017
... id="C12">丝状真菌次级代谢与其基因簇所处的表观遗传状态有着密切的关系.通过表观遗传操作能大范围改变代谢物谱,以及激活隐性次级代谢物的产生[37].表观遗传调控是一种基于核酸修饰的调控方式,主要包括DNA甲基化、组蛋白甲基化、乙酰化、磷酸化和泛素化等.其中,组蛋白甲基化是最稳定的修饰方式,因此最适合作为稳定的表观遗传信息.而乙酰化修饰则具有较高的动态性,同时磷酸化和泛素化修饰相对不太稳定,因此它们更为灵活地影响染色质的结构和功能.组蛋白遗传修饰之间相互关联并共同控制着基因的表达,从而最终影响丝状真菌次级代谢物的合成.其中,组蛋白甲基化和乙酰化研究的最为透彻.LaeA作为组蛋白甲基转移酶,通过调控组蛋白和改变染色体结构来调控次级代谢产物的合成.通常组蛋白甲基转移酶和组蛋白去乙酰化酶(HDAC)能够阻遏次级代谢物的合成.浙江大学毛旭明、中国医学科学院药物研究所胡友财课题组以及美国加州大学唐奕课题组合作研究发现,敲除齿梗孢霉(Calcarisporium arbuscula)中组蛋白去乙酰化酶使得菌体中超过75%的次级代谢基因簇表达水平上升,从该突变株中分离的10种化合物中4种具有新结构[38],该工作提示可以通过表观遗传的改变快速同时激活多个隐性基因簇,为基因组挖掘提供了强大的工具.朱旭东课题组发现小孢拟盘多毛孢组蛋白乙酰转移酶基因hat1正调控次级代谢和分生孢子的形成,同时也控制细胞壁的完整性[39].中国农业科学院植物保护研究所冯洁课题组发现敲除组蛋白乙酰转移酶基因FgSAS3和FgGCN5增加了禾谷镰刀菌对渗透压和活性氧(ROS)的敏感性,FgSAS3敲除株中分生孢子的量和子囊壳的形成减少,而FgGCN5敲除株则完全不能形成分生孢子和子囊壳.同时发现FgSAS3和FgGCN5是禾谷镰刀菌感染小麦穗和番茄果实的重要致病因子.FgSAS3和FgGCN敲除株不再产生DON,这一结果与敲除株中参与DON合成的关键基因的转录明显下降相一致[40].福建农林大学汪世华课题组发现黄曲霉中组蛋白乙酰转移酶基因AflgcnE的缺失抑制了黄曲霉的生长,降低了细胞表面的疏水性,同时使得菌体不能产生无性孢子和菌核,并导致黄曲霉毒素合成相关基因表达量急剧下降,黄曲霉毒素不再产生[41].尹文兵课题组发现敲除无花果拟盘多毛孢(Pestalotiopsis fici)中组蛋白甲基转移酶基因PfcclA和组蛋白去乙酰化酶基因PfhdaA激活了15种新的聚酮化合物的产生,同时提高了11种大环内酯类抗生素的产量[42].另外罗氏绿僵菌(Metarhizium robertsii)中hat1的敲除激活了孤儿次级代谢物生物合成基因的表达,在该敲除株中分离得到了11种新化合物[43].近年来的研究发现,细菌和真菌中还普遍存在组蛋白赖氨酸的琥珀酰化修饰,并且已经证明其在细胞分化、中心代谢等重要生命活动中发挥着重要作用,尤其是西南大学谢建平课题组对结核分枝杆菌琥珀酰化与其致病性关系进行了详细的蛋白组学研究,为结核病的诊断和防治提供了科学依据[44].然而,截至目前为止还没有在丝状真菌中发现琥珀酰化与次级代谢产物合成之间关系的报道.总之,上述研究证明表观遗传调控是发掘新化合物便捷和有效的策略之一. ...
结核分枝杆菌表观遗传与耐药新机理研究 [学位论文]
1
2017
... id="C12">丝状真菌次级代谢与其基因簇所处的表观遗传状态有着密切的关系.通过表观遗传操作能大范围改变代谢物谱,以及激活隐性次级代谢物的产生[37].表观遗传调控是一种基于核酸修饰的调控方式,主要包括DNA甲基化、组蛋白甲基化、乙酰化、磷酸化和泛素化等.其中,组蛋白甲基化是最稳定的修饰方式,因此最适合作为稳定的表观遗传信息.而乙酰化修饰则具有较高的动态性,同时磷酸化和泛素化修饰相对不太稳定,因此它们更为灵活地影响染色质的结构和功能.组蛋白遗传修饰之间相互关联并共同控制着基因的表达,从而最终影响丝状真菌次级代谢物的合成.其中,组蛋白甲基化和乙酰化研究的最为透彻.LaeA作为组蛋白甲基转移酶,通过调控组蛋白和改变染色体结构来调控次级代谢产物的合成.通常组蛋白甲基转移酶和组蛋白去乙酰化酶(HDAC)能够阻遏次级代谢物的合成.浙江大学毛旭明、中国医学科学院药物研究所胡友财课题组以及美国加州大学唐奕课题组合作研究发现,敲除齿梗孢霉(Calcarisporium arbuscula)中组蛋白去乙酰化酶使得菌体中超过75%的次级代谢基因簇表达水平上升,从该突变株中分离的10种化合物中4种具有新结构[38],该工作提示可以通过表观遗传的改变快速同时激活多个隐性基因簇,为基因组挖掘提供了强大的工具.朱旭东课题组发现小孢拟盘多毛孢组蛋白乙酰转移酶基因hat1正调控次级代谢和分生孢子的形成,同时也控制细胞壁的完整性[39].中国农业科学院植物保护研究所冯洁课题组发现敲除组蛋白乙酰转移酶基因FgSAS3和FgGCN5增加了禾谷镰刀菌对渗透压和活性氧(ROS)的敏感性,FgSAS3敲除株中分生孢子的量和子囊壳的形成减少,而FgGCN5敲除株则完全不能形成分生孢子和子囊壳.同时发现FgSAS3和FgGCN5是禾谷镰刀菌感染小麦穗和番茄果实的重要致病因子.FgSAS3和FgGCN敲除株不再产生DON,这一结果与敲除株中参与DON合成的关键基因的转录明显下降相一致[40].福建农林大学汪世华课题组发现黄曲霉中组蛋白乙酰转移酶基因AflgcnE的缺失抑制了黄曲霉的生长,降低了细胞表面的疏水性,同时使得菌体不能产生无性孢子和菌核,并导致黄曲霉毒素合成相关基因表达量急剧下降,黄曲霉毒素不再产生[41].尹文兵课题组发现敲除无花果拟盘多毛孢(Pestalotiopsis fici)中组蛋白甲基转移酶基因PfcclA和组蛋白去乙酰化酶基因PfhdaA激活了15种新的聚酮化合物的产生,同时提高了11种大环内酯类抗生素的产量[42].另外罗氏绿僵菌(Metarhizium robertsii)中hat1的敲除激活了孤儿次级代谢物生物合成基因的表达,在该敲除株中分离得到了11种新化合物[43].近年来的研究发现,细菌和真菌中还普遍存在组蛋白赖氨酸的琥珀酰化修饰,并且已经证明其在细胞分化、中心代谢等重要生命活动中发挥着重要作用,尤其是西南大学谢建平课题组对结核分枝杆菌琥珀酰化与其致病性关系进行了详细的蛋白组学研究,为结核病的诊断和防治提供了科学依据[44].然而,截至目前为止还没有在丝状真菌中发现琥珀酰化与次级代谢产物合成之间关系的报道.总之,上述研究证明表观遗传调控是发掘新化合物便捷和有效的策略之一. ...
Transcriptomic profiling of Aspergillus flavus in response to 5-azacytidine
1
2013
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
DNA methyltransferase inhibitor dramatically alters the secondary metabolism of Pestalotiopsis microspora
1
2017
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
Induced production of new diterpenoids in the fungus Penicillium funiculosum
1
2014
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
Epigenetic modifiers alter the secondary metabolite composition of a plant endophytic fungus, Pestalotiopsis crassiuscula obtained from the leaves of Fragaria chiloensis
1
2014
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
Identification and biological evaluation of secondary metabolites from marine derived fungi- Aspergillus sp
1
2017
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
Eremophilane sesquiterpenes from a deep marine-derived fungus, Aspergillus sp. SCSIOW2, cultivated in the presence of epigenetic modifying agents
1
2016
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
DNA methyltransferase inhibitor induced fungal biosynthetic products: Diethylene glycol phthalate ester oligomers from the marine-derived fungus Cochliobolus lunatus
1
2016
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
Brominated resorcylic acid lactones from the marine- derived fungus Cochliobolus lunatus induced by histone deacetylase inhibitors
1
2014
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
Fermentation optimization for the gorgonian coral-derived fungus Curvularia lunatato produce cytochalasin B and the study of adding chemical epigenetic modification in fermentation
1
2016
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
Discovery of a new biphenyl derivative by epigenetic manipulation of marine-derived fungus Aspergillus versicolor
1
2018
... id="C13">通过直接阻断或高表达表观遗传相关基因能够提高抗生素产量和发现新型次级代谢物.然而,由于很多丝状真菌缺乏有效的遗传操作体系,无法采用遗传手段来研究表观遗传与次级代谢的关系.近年来,一系列特异地抑制表观遗传修饰酶活性的化学试剂被发现,通过这些化学试剂来调控次级代谢,最终形成了一门新的学科,即化学表观遗传学.中山大学贺竹梅课题组利用DNA甲基转移酶抑制剂5-杂氮胞苷(5-AC)处理黄曲霉,发现5-AC阻遏了黄曲霉的形态分化和黄曲霉毒素的生物合成,随后RNA-seq结果表明VeA的表达受到5-AC的强烈抑制,推测5-AC抑制了VelB-VeA-LaeA复合物的形成以及羧肽酶的活性,从而降低了黄曲霉毒素的产量[45].重庆大学杨小龙课题组发现,5-AC能够诱 导小孢拟盘多毛孢产生新化合物4′-formamidophenyl-5-methoxybenzoate和7种已知聚酮化合物[46].绳状青霉菌(P. funiculosum)经过5-AC处理诱导后产生两种新的prenyleudesmane二萜类化合物[47].在P. crassiuscula中加入5-AC,导致该菌产生了包括coumarin在内的4种新化合物[48].上述证据表明,5-AC作为DNA甲基转移酶抑制剂能够激活隐性基因簇并获得新型代谢物.通过化学试剂抑制组蛋白的去乙酰化,有可能提高组蛋白乙酰化水平,促进基因的表达.深圳大学王力彦课题组用组蛋白去乙酰化酶抑制剂SAHA和5-AC共同处理两株深海来源的曲霉,其中一株产生了1种新型的二苯醚-O-糖苷和7种已知化合物[49],另外一株产生了3种新的倍半萜类化合物和1种已知化合物[50].中国海洋大学王长云课题组利用5-AC和组蛋白去乙酰化酶抑制剂处理Cochliobolus lunatus,激活了包括cochphthesters A-G在内的9种新型化合物[51, 52].他们在Curvularia lunata中加入组蛋白去乙酰化酶抑制剂丁酸钠后,激活了4种化合物的产生[53].宁波大学何山课题组利用SAHA处理杂色曲霉,激活了新的联苯类化合物versiperol A以及已知化合物2,4-dimethoxyphenol和diorcino的产生[54]. ...
Disruption of a glutathione reductase encoding gene in Acremonium chrysogenum leads to reduction of its growth, cephalosporin production and antioxidative ability which is recovered by exogenous methionine
1
2012
... id="C15">丝状真菌中氧化胁迫往往与其次级代谢息息相关,而谷胱甘肽和硫氧还蛋白还原酶系统是丝状真菌应对外来氧化胁迫的重要方式.本课题组发现敲除顶头孢霉谷胱甘肽还原酶基因glrA或硫氧还蛋白还原酶基因ActrxR1都能延迟孢子萌发和菌丝生长;与ActrxR1不同,glrA的缺失还增加了菌体对过氧化氢的敏感度;glrA的缺失完全阻断了头孢菌素的合成,但这种缺陷可以被外源添加甲硫氨酸所弥补,而在ActrxR1突变株中头孢菌素的产量不但没有减少,反而增加了一倍[55, 56].在维持体内氧化还原平衡方面,glrA和ActrxR1的作用也完全不同(图1).推测AcTrxR1用于消除体内低浓度的ROS,而低浓度的ROS可以刺激次级代谢物的合成;相反,高浓度的ROS则对细胞有毒,甚至可以导致细胞死亡,同时也不利于次级代谢物的产生,而GlrA则用于应对体内产生的高浓度ROS.bZIP类调控因子在丝状真菌中保守存在,它在响应环境信号过程中发挥着重要作用.某些bZIP转录因子在调控ROS产生的同时,也调控了次级代谢物的合成.尹文兵课题组发现bZIP类调控因子编码基因PfbzipA的缺失增强了无花果拟盘多毛孢对叔丁基过氧化氢、偶氮二甲苯、甲基萘醌亚硫酸氢钠的耐性,但对过氧化氢却更加敏感.同时PfbzipA的敲除也改变了菌体产生次级代谢物的能力[57]. ...
The thioredoxinreductase-encoding gene ActrxR1 is involved in the cephalosporin C production of Acremonium chrysogenum in methionine-supplemented medium
1
2013
... id="C15">丝状真菌中氧化胁迫往往与其次级代谢息息相关,而谷胱甘肽和硫氧还蛋白还原酶系统是丝状真菌应对外来氧化胁迫的重要方式.本课题组发现敲除顶头孢霉谷胱甘肽还原酶基因glrA或硫氧还蛋白还原酶基因ActrxR1都能延迟孢子萌发和菌丝生长;与ActrxR1不同,glrA的缺失还增加了菌体对过氧化氢的敏感度;glrA的缺失完全阻断了头孢菌素的合成,但这种缺陷可以被外源添加甲硫氨酸所弥补,而在ActrxR1突变株中头孢菌素的产量不但没有减少,反而增加了一倍[55, 56].在维持体内氧化还原平衡方面,glrA和ActrxR1的作用也完全不同(图1).推测AcTrxR1用于消除体内低浓度的ROS,而低浓度的ROS可以刺激次级代谢物的合成;相反,高浓度的ROS则对细胞有毒,甚至可以导致细胞死亡,同时也不利于次级代谢物的产生,而GlrA则用于应对体内产生的高浓度ROS.bZIP类调控因子在丝状真菌中保守存在,它在响应环境信号过程中发挥着重要作用.某些bZIP转录因子在调控ROS产生的同时,也调控了次级代谢物的合成.尹文兵课题组发现bZIP类调控因子编码基因PfbzipA的缺失增强了无花果拟盘多毛孢对叔丁基过氧化氢、偶氮二甲苯、甲基萘醌亚硫酸氢钠的耐性,但对过氧化氢却更加敏感.同时PfbzipA的敲除也改变了菌体产生次级代谢物的能力[57]. ...
The bZIP transcription factor PfZipA regulates secondary metabolism and oxidative stress response in the plant endophytic fungus Pestalotiopsis fici
1
2015
... id="C15">丝状真菌中氧化胁迫往往与其次级代谢息息相关,而谷胱甘肽和硫氧还蛋白还原酶系统是丝状真菌应对外来氧化胁迫的重要方式.本课题组发现敲除顶头孢霉谷胱甘肽还原酶基因glrA或硫氧还蛋白还原酶基因ActrxR1都能延迟孢子萌发和菌丝生长;与ActrxR1不同,glrA的缺失还增加了菌体对过氧化氢的敏感度;glrA的缺失完全阻断了头孢菌素的合成,但这种缺陷可以被外源添加甲硫氨酸所弥补,而在ActrxR1突变株中头孢菌素的产量不但没有减少,反而增加了一倍[55, 56].在维持体内氧化还原平衡方面,glrA和ActrxR1的作用也完全不同(图1).推测AcTrxR1用于消除体内低浓度的ROS,而低浓度的ROS可以刺激次级代谢物的合成;相反,高浓度的ROS则对细胞有毒,甚至可以导致细胞死亡,同时也不利于次级代谢物的产生,而GlrA则用于应对体内产生的高浓度ROS.bZIP类调控因子在丝状真菌中保守存在,它在响应环境信号过程中发挥着重要作用.某些bZIP转录因子在调控ROS产生的同时,也调控了次级代谢物的合成.尹文兵课题组发现bZIP类调控因子编码基因PfbzipA的缺失增强了无花果拟盘多毛孢对叔丁基过氧化氢、偶氮二甲苯、甲基萘醌亚硫酸氢钠的耐性,但对过氧化氢却更加敏感.同时PfbzipA的敲除也改变了菌体产生次级代谢物的能力[57]. ...
Disruption of the nitrogen regulatory gene AcareA in Acremonium chrysogenum leads to reduction of cephalosporin production and repression of nitrogen metabolism
1
2013
... id="C17">丝状真菌次级代谢还受到氮源的调控.AREA是含有锌指结构域的GATA类调控蛋白,通过与氮源利用相关基因启动子区的结合参与丝状真菌氮源调控.AREA还能直接调控次级代谢物生物合成基因的转录.本课题组发现顶头孢霉中氮源调控基因AcareA和AcareB除了调控氮源代谢,还能够调控头孢菌素的生物合成.AcAREA通过与头孢菌素生物合成关键基因pcbAB-pcbC双向启动子区的结合直接调控pcbAB和pcbC的转录,间接促进cefD2、cefEF和cefG的转录[58].而AcAREB则通过与pcbAB- pcbC、cefD1-cefD2以及cefEF-cefG的双向启动子区结合直接正调控头孢菌素的产生[59]. ...
A GATA-type transcription factor AcAREB for nitrogen metabolism is involved in regulation of cephalosporin biosynthesis in Acremonium chrysogenum
1
2017
... id="C17">丝状真菌次级代谢还受到氮源的调控.AREA是含有锌指结构域的GATA类调控蛋白,通过与氮源利用相关基因启动子区的结合参与丝状真菌氮源调控.AREA还能直接调控次级代谢物生物合成基因的转录.本课题组发现顶头孢霉中氮源调控基因AcareA和AcareB除了调控氮源代谢,还能够调控头孢菌素的生物合成.AcAREA通过与头孢菌素生物合成关键基因pcbAB-pcbC双向启动子区的结合直接调控pcbAB和pcbC的转录,间接促进cefD2、cefEF和cefG的转录[58].而AcAREB则通过与pcbAB- pcbC、cefD1-cefD2以及cefEF-cefG的双向启动子区结合直接正调控头孢菌素的产生[59]. ...
The pH-responsive transcription factor PacC regulates mycelial growth, fruiting body development, and ganoderic acid biosynthesis in Ganoderma lucidum
1
2016
... id="C18">应答调控pH变化的转录因子PacC也与次级代谢紧密相关.南京农业大学赵明文课题组发现赤芝(Ganoderma lucidum)中GlPacC负调控灵芝酸的合成[60].汪世华课题组发现敲除碳代谢阻遏基因creA导致黄曲霉生长缺陷,分生孢子量减少,黄曲霉毒素产量明显降低[61],这是碳源对次级代谢物生物合成调控的典型特征.中国科学科院华南植物园蒋跃明课题组发现橘青霉中Zn(II)2Cys6类转录因子、LaeA以及PacC在以葡萄糖为碳源的培养基中的表达量均高于以蔗糖为碳源的培养基,而GATA类转录因子的表达在这两种培养基中的表现正好相反[62].温度对次级代谢物的生物合成也有重要的影响,天津科技大学王昌禄课题组研究发现,红曲霉菌在变温培养(30~25℃)时较恒温培养(30℃)时monacolin K的产量提高了16倍.蛋白质组学结果表明:变温培养增加了次级代谢物生物合成途径相关基因的表达,同时却阻遏了与菌丝生长相关的基因表达[63].此外,VeA作为参与光照调控的重要调控因子也是连接VelB和LaeA形成复合体的重要桥梁.多种微生物共培养可以刺激产生新的次级代谢物.华南农业大学李春远课题组研究发现,两株红树林真菌Phomopsis sp. K38和Alternariasp. E33共培养后刺激了环肽化合物[64]、二酰亚胺化合物[65]和四肽化合物[66]的产生.同时海洋真菌Emericella sp.与放线菌Salinispora arenicola共培养刺激了emericellamides的大量产生[67].类似的现象还很多,这些次级代谢产物在微生物间的通讯中也扮演着重要的角色.因此,多种微生物的共培养为隐形次级代谢物的激活提供了新的策略. ...
Carbon catabolite repression gene creA regulates morphology, aflatoxin biosynthesis and virulence in Aspergillus flavus
1
2018
... id="C18">应答调控pH变化的转录因子PacC也与次级代谢紧密相关.南京农业大学赵明文课题组发现赤芝(Ganoderma lucidum)中GlPacC负调控灵芝酸的合成[60].汪世华课题组发现敲除碳代谢阻遏基因creA导致黄曲霉生长缺陷,分生孢子量减少,黄曲霉毒素产量明显降低[61],这是碳源对次级代谢物生物合成调控的典型特征.中国科学科院华南植物园蒋跃明课题组发现橘青霉中Zn(II)2Cys6类转录因子、LaeA以及PacC在以葡萄糖为碳源的培养基中的表达量均高于以蔗糖为碳源的培养基,而GATA类转录因子的表达在这两种培养基中的表现正好相反[62].温度对次级代谢物的生物合成也有重要的影响,天津科技大学王昌禄课题组研究发现,红曲霉菌在变温培养(30~25℃)时较恒温培养(30℃)时monacolin K的产量提高了16倍.蛋白质组学结果表明:变温培养增加了次级代谢物生物合成途径相关基因的表达,同时却阻遏了与菌丝生长相关的基因表达[63].此外,VeA作为参与光照调控的重要调控因子也是连接VelB和LaeA形成复合体的重要桥梁.多种微生物共培养可以刺激产生新的次级代谢物.华南农业大学李春远课题组研究发现,两株红树林真菌Phomopsis sp. K38和Alternariasp. E33共培养后刺激了环肽化合物[64]、二酰亚胺化合物[65]和四肽化合物[66]的产生.同时海洋真菌Emericella sp.与放线菌Salinispora arenicola共培养刺激了emericellamides的大量产生[67].类似的现象还很多,这些次级代谢产物在微生物间的通讯中也扮演着重要的角色.因此,多种微生物的共培养为隐形次级代谢物的激活提供了新的策略. ...
Comparative transcriptome analysis of Penicillium citrinum cultured with different carbon sources identifies genes involved in citrinin biosynthesis
1
2017
... id="C18">应答调控pH变化的转录因子PacC也与次级代谢紧密相关.南京农业大学赵明文课题组发现赤芝(Ganoderma lucidum)中GlPacC负调控灵芝酸的合成[60].汪世华课题组发现敲除碳代谢阻遏基因creA导致黄曲霉生长缺陷,分生孢子量减少,黄曲霉毒素产量明显降低[61],这是碳源对次级代谢物生物合成调控的典型特征.中国科学科院华南植物园蒋跃明课题组发现橘青霉中Zn(II)2Cys6类转录因子、LaeA以及PacC在以葡萄糖为碳源的培养基中的表达量均高于以蔗糖为碳源的培养基,而GATA类转录因子的表达在这两种培养基中的表现正好相反[62].温度对次级代谢物的生物合成也有重要的影响,天津科技大学王昌禄课题组研究发现,红曲霉菌在变温培养(30~25℃)时较恒温培养(30℃)时monacolin K的产量提高了16倍.蛋白质组学结果表明:变温培养增加了次级代谢物生物合成途径相关基因的表达,同时却阻遏了与菌丝生长相关的基因表达[63].此外,VeA作为参与光照调控的重要调控因子也是连接VelB和LaeA形成复合体的重要桥梁.多种微生物共培养可以刺激产生新的次级代谢物.华南农业大学李春远课题组研究发现,两株红树林真菌Phomopsis sp. K38和Alternariasp. E33共培养后刺激了环肽化合物[64]、二酰亚胺化合物[65]和四肽化合物[66]的产生.同时海洋真菌Emericella sp.与放线菌Salinispora arenicola共培养刺激了emericellamides的大量产生[67].类似的现象还很多,这些次级代谢产物在微生物间的通讯中也扮演着重要的角色.因此,多种微生物的共培养为隐形次级代谢物的激活提供了新的策略. ...
Effect of temperature-shift and temperature-constant cultivation on the Monacolin K biosynthetic gene cluster expression in Monascus sp
1
2017
... id="C18">应答调控pH变化的转录因子PacC也与次级代谢紧密相关.南京农业大学赵明文课题组发现赤芝(Ganoderma lucidum)中GlPacC负调控灵芝酸的合成[60].汪世华课题组发现敲除碳代谢阻遏基因creA导致黄曲霉生长缺陷,分生孢子量减少,黄曲霉毒素产量明显降低[61],这是碳源对次级代谢物生物合成调控的典型特征.中国科学科院华南植物园蒋跃明课题组发现橘青霉中Zn(II)2Cys6类转录因子、LaeA以及PacC在以葡萄糖为碳源的培养基中的表达量均高于以蔗糖为碳源的培养基,而GATA类转录因子的表达在这两种培养基中的表现正好相反[62].温度对次级代谢物的生物合成也有重要的影响,天津科技大学王昌禄课题组研究发现,红曲霉菌在变温培养(30~25℃)时较恒温培养(30℃)时monacolin K的产量提高了16倍.蛋白质组学结果表明:变温培养增加了次级代谢物生物合成途径相关基因的表达,同时却阻遏了与菌丝生长相关的基因表达[63].此外,VeA作为参与光照调控的重要调控因子也是连接VelB和LaeA形成复合体的重要桥梁.多种微生物共培养可以刺激产生新的次级代谢物.华南农业大学李春远课题组研究发现,两株红树林真菌Phomopsis sp. K38和Alternariasp. E33共培养后刺激了环肽化合物[64]、二酰亚胺化合物[65]和四肽化合物[66]的产生.同时海洋真菌Emericella sp.与放线菌Salinispora arenicola共培养刺激了emericellamides的大量产生[67].类似的现象还很多,这些次级代谢产物在微生物间的通讯中也扮演着重要的角色.因此,多种微生物的共培养为隐形次级代谢物的激活提供了新的策略. ...
A new cyclopeptide with antifungal activity from the co-culture broth of two marine mangrove fungi
1
2014
... id="C18">应答调控pH变化的转录因子PacC也与次级代谢紧密相关.南京农业大学赵明文课题组发现赤芝(Ganoderma lucidum)中GlPacC负调控灵芝酸的合成[60].汪世华课题组发现敲除碳代谢阻遏基因creA导致黄曲霉生长缺陷,分生孢子量减少,黄曲霉毒素产量明显降低[61],这是碳源对次级代谢物生物合成调控的典型特征.中国科学科院华南植物园蒋跃明课题组发现橘青霉中Zn(II)2Cys6类转录因子、LaeA以及PacC在以葡萄糖为碳源的培养基中的表达量均高于以蔗糖为碳源的培养基,而GATA类转录因子的表达在这两种培养基中的表现正好相反[62].温度对次级代谢物的生物合成也有重要的影响,天津科技大学王昌禄课题组研究发现,红曲霉菌在变温培养(30~25℃)时较恒温培养(30℃)时monacolin K的产量提高了16倍.蛋白质组学结果表明:变温培养增加了次级代谢物生物合成途径相关基因的表达,同时却阻遏了与菌丝生长相关的基因表达[63].此外,VeA作为参与光照调控的重要调控因子也是连接VelB和LaeA形成复合体的重要桥梁.多种微生物共培养可以刺激产生新的次级代谢物.华南农业大学李春远课题组研究发现,两株红树林真菌Phomopsis sp. K38和Alternariasp. E33共培养后刺激了环肽化合物[64]、二酰亚胺化合物[65]和四肽化合物[66]的产生.同时海洋真菌Emericella sp.与放线菌Salinispora arenicola共培养刺激了emericellamides的大量产生[67].类似的现象还很多,这些次级代谢产物在微生物间的通讯中也扮演着重要的角色.因此,多种微生物的共培养为隐形次级代谢物的激活提供了新的策略. ...
A new diimide derivative from the co-culture broth of two mangrove fungi (strain no. E33 and K38)
1
2010
... id="C18">应答调控pH变化的转录因子PacC也与次级代谢紧密相关.南京农业大学赵明文课题组发现赤芝(Ganoderma lucidum)中GlPacC负调控灵芝酸的合成[60].汪世华课题组发现敲除碳代谢阻遏基因creA导致黄曲霉生长缺陷,分生孢子量减少,黄曲霉毒素产量明显降低[61],这是碳源对次级代谢物生物合成调控的典型特征.中国科学科院华南植物园蒋跃明课题组发现橘青霉中Zn(II)2Cys6类转录因子、LaeA以及PacC在以葡萄糖为碳源的培养基中的表达量均高于以蔗糖为碳源的培养基,而GATA类转录因子的表达在这两种培养基中的表现正好相反[62].温度对次级代谢物的生物合成也有重要的影响,天津科技大学王昌禄课题组研究发现,红曲霉菌在变温培养(30~25℃)时较恒温培养(30℃)时monacolin K的产量提高了16倍.蛋白质组学结果表明:变温培养增加了次级代谢物生物合成途径相关基因的表达,同时却阻遏了与菌丝生长相关的基因表达[63].此外,VeA作为参与光照调控的重要调控因子也是连接VelB和LaeA形成复合体的重要桥梁.多种微生物共培养可以刺激产生新的次级代谢物.华南农业大学李春远课题组研究发现,两株红树林真菌Phomopsis sp. K38和Alternariasp. E33共培养后刺激了环肽化合物[64]、二酰亚胺化合物[65]和四肽化合物[66]的产生.同时海洋真菌Emericella sp.与放线菌Salinispora arenicola共培养刺激了emericellamides的大量产生[67].类似的现象还很多,这些次级代谢产物在微生物间的通讯中也扮演着重要的角色.因此,多种微生物的共培养为隐形次级代谢物的激活提供了新的策略. ...
Two new cyclopeptides from the co-culture broth of two marine mangrove fungi and their antifungal activity
1
2014
... id="C18">应答调控pH变化的转录因子PacC也与次级代谢紧密相关.南京农业大学赵明文课题组发现赤芝(Ganoderma lucidum)中GlPacC负调控灵芝酸的合成[60].汪世华课题组发现敲除碳代谢阻遏基因creA导致黄曲霉生长缺陷,分生孢子量减少,黄曲霉毒素产量明显降低[61],这是碳源对次级代谢物生物合成调控的典型特征.中国科学科院华南植物园蒋跃明课题组发现橘青霉中Zn(II)2Cys6类转录因子、LaeA以及PacC在以葡萄糖为碳源的培养基中的表达量均高于以蔗糖为碳源的培养基,而GATA类转录因子的表达在这两种培养基中的表现正好相反[62].温度对次级代谢物的生物合成也有重要的影响,天津科技大学王昌禄课题组研究发现,红曲霉菌在变温培养(30~25℃)时较恒温培养(30℃)时monacolin K的产量提高了16倍.蛋白质组学结果表明:变温培养增加了次级代谢物生物合成途径相关基因的表达,同时却阻遏了与菌丝生长相关的基因表达[63].此外,VeA作为参与光照调控的重要调控因子也是连接VelB和LaeA形成复合体的重要桥梁.多种微生物共培养可以刺激产生新的次级代谢物.华南农业大学李春远课题组研究发现,两株红树林真菌Phomopsis sp. K38和Alternariasp. E33共培养后刺激了环肽化合物[64]、二酰亚胺化合物[65]和四肽化合物[66]的产生.同时海洋真菌Emericella sp.与放线菌Salinispora arenicola共培养刺激了emericellamides的大量产生[67].类似的现象还很多,这些次级代谢产物在微生物间的通讯中也扮演着重要的角色.因此,多种微生物的共培养为隐形次级代谢物的激活提供了新的策略. ...
Induced production of emericellamides A and B from the marine- derived fungus Emericella sp. in competing co-culture
1
2007
... id="C18">应答调控pH变化的转录因子PacC也与次级代谢紧密相关.南京农业大学赵明文课题组发现赤芝(Ganoderma lucidum)中GlPacC负调控灵芝酸的合成[60].汪世华课题组发现敲除碳代谢阻遏基因creA导致黄曲霉生长缺陷,分生孢子量减少,黄曲霉毒素产量明显降低[61],这是碳源对次级代谢物生物合成调控的典型特征.中国科学科院华南植物园蒋跃明课题组发现橘青霉中Zn(II)2Cys6类转录因子、LaeA以及PacC在以葡萄糖为碳源的培养基中的表达量均高于以蔗糖为碳源的培养基,而GATA类转录因子的表达在这两种培养基中的表现正好相反[62].温度对次级代谢物的生物合成也有重要的影响,天津科技大学王昌禄课题组研究发现,红曲霉菌在变温培养(30~25℃)时较恒温培养(30℃)时monacolin K的产量提高了16倍.蛋白质组学结果表明:变温培养增加了次级代谢物生物合成途径相关基因的表达,同时却阻遏了与菌丝生长相关的基因表达[63].此外,VeA作为参与光照调控的重要调控因子也是连接VelB和LaeA形成复合体的重要桥梁.多种微生物共培养可以刺激产生新的次级代谢物.华南农业大学李春远课题组研究发现,两株红树林真菌Phomopsis sp. K38和Alternariasp. E33共培养后刺激了环肽化合物[64]、二酰亚胺化合物[65]和四肽化合物[66]的产生.同时海洋真菌Emericella sp.与放线菌Salinispora arenicola共培养刺激了emericellamides的大量产生[67].类似的现象还很多,这些次级代谢产物在微生物间的通讯中也扮演着重要的角色.因此,多种微生物的共培养为隐形次级代谢物的激活提供了新的策略. ...
顶头孢霉高低产菌初级代谢的比较及CPC甲氧基化的探索 [学位论文]
1
2016
... id="C19">次级代谢与初级代谢息息相关,初级代谢为次级代谢物的合成提供前体和辅因子.次级代谢的前体可以是中心碳代谢的中间体,也可以是参与合成代谢反应的细胞组成元件.通常认为次级代谢从中心碳代谢获取代谢物的能力依赖于代谢物在细胞内的浓度以及次级代谢途径第一个反应酶的活力.通向中心碳代谢的代谢流通常比通向次级代谢合成的代谢流高很多,当通向次级代谢物合成的代谢流增加时,会显著地影响到中心碳代谢代谢物的浓度和次级代谢途径第一个酶转化前体的能力,此时可以通过提高次级代谢物生物合成基因的表达而提高次级代谢物的产量.然而,当前体是细胞组成元件时,将不能通过增加生物合成基因或者调控基因的表达而改变次级代谢流.初级代谢除了为次级代谢提供合成前体外,还可以为其提供辅因子,例如在青霉素和头孢菌素等β-内酰胺类抗生素合成过程中需要大量的NADPH作为还原力.为了满足青霉素合成对NADPH的需求,可以增加戊糖磷酸途径中的代谢流.通过比对分析青霉素高产菌株和低产菌株中心碳代谢流发现,高产菌株通过戊糖磷酸途径的代谢流高于低产菌株,这一结果与青霉素合成需要大量NADPH恰恰一致[68].推测丝状真菌对前体和辅因子需求量的增加也是限制抗生素产量提高的重要因素,因此可以通过改造丝状真菌初级代谢途径来增加次级代谢物的产量. ...
顶头孢霉高低产菌初级代谢的比较及CPC甲氧基化的探索 [学位论文]
1
2016
... id="C19">次级代谢与初级代谢息息相关,初级代谢为次级代谢物的合成提供前体和辅因子.次级代谢的前体可以是中心碳代谢的中间体,也可以是参与合成代谢反应的细胞组成元件.通常认为次级代谢从中心碳代谢获取代谢物的能力依赖于代谢物在细胞内的浓度以及次级代谢途径第一个反应酶的活力.通向中心碳代谢的代谢流通常比通向次级代谢合成的代谢流高很多,当通向次级代谢物合成的代谢流增加时,会显著地影响到中心碳代谢代谢物的浓度和次级代谢途径第一个酶转化前体的能力,此时可以通过提高次级代谢物生物合成基因的表达而提高次级代谢物的产量.然而,当前体是细胞组成元件时,将不能通过增加生物合成基因或者调控基因的表达而改变次级代谢流.初级代谢除了为次级代谢提供合成前体外,还可以为其提供辅因子,例如在青霉素和头孢菌素等β-内酰胺类抗生素合成过程中需要大量的NADPH作为还原力.为了满足青霉素合成对NADPH的需求,可以增加戊糖磷酸途径中的代谢流.通过比对分析青霉素高产菌株和低产菌株中心碳代谢流发现,高产菌株通过戊糖磷酸途径的代谢流高于低产菌株,这一结果与青霉素合成需要大量NADPH恰恰一致[68].推测丝状真菌对前体和辅因子需求量的增加也是限制抗生素产量提高的重要因素,因此可以通过改造丝状真菌初级代谢途径来增加次级代谢物的产量. ...
Relationship between secondary metabolism and fungal development
1
2002
... id="C20">丝状真菌次级代谢物什么时间产生是一个重要的科学问题,普遍认为它产生于生命周期特定的时间点和特定的环境条件下.次级代谢物合成的开启也被认为是初级代谢过程的结束.初级代谢向次级代谢的转换往往与营养耗尽、光照缺少、生殖结构发育、pH变化等密切相关.研究表明构巢曲霉和黄曲霉中次级代谢物合成的开启与有性子实体的形成和菌核的产生有很强的关联性,分生孢子的产生与参与孢子色素合成相关基因的表达也有很强的相关性[69].初级代谢向次级代谢转换的过程中往往伴随着信号分子的产生,而信号分子对次级代谢物的产生具有诱导作用.如群感效应分子在链霉菌次级代谢物合成的开启中起到决定性的作用一样,群感效应分子与丝状真菌的形态分化和次级代谢物的产生也具有相关性.如构巢曲霉和黄曲霉中的群感效应分子氧脂素不仅在调节有性孢子和无性孢子的比例中起关键作用,同时还影响柄曲霉素和黄曲霉毒素的产生[70].除了信号分子诱导之外,环境压力胁迫也是次级代谢产物开启的重要因素,例如委内瑞拉链霉菌中杰多霉素B的产生需要噬菌体的入侵或者乙醇的诱导[71].丝状真菌中也存在类似的现象,如黄曲霉中活性氧的产生或者体外添加氧化物能够诱导黄曲霉毒素的产生[72].然而关于群感效应分子和胁迫诱导丝状真菌次级代谢物产生的作用机制尚不清楚,这也是未来丝状真菌次级代谢研究需要加强的地方. ...
Oxygenase coordination is required for morphological transition and the host-fungus interaction of Aspergillus flavus
1
2009
... id="C20">丝状真菌次级代谢物什么时间产生是一个重要的科学问题,普遍认为它产生于生命周期特定的时间点和特定的环境条件下.次级代谢物合成的开启也被认为是初级代谢过程的结束.初级代谢向次级代谢的转换往往与营养耗尽、光照缺少、生殖结构发育、pH变化等密切相关.研究表明构巢曲霉和黄曲霉中次级代谢物合成的开启与有性子实体的形成和菌核的产生有很强的关联性,分生孢子的产生与参与孢子色素合成相关基因的表达也有很强的相关性[69].初级代谢向次级代谢转换的过程中往往伴随着信号分子的产生,而信号分子对次级代谢物的产生具有诱导作用.如群感效应分子在链霉菌次级代谢物合成的开启中起到决定性的作用一样,群感效应分子与丝状真菌的形态分化和次级代谢物的产生也具有相关性.如构巢曲霉和黄曲霉中的群感效应分子氧脂素不仅在调节有性孢子和无性孢子的比例中起关键作用,同时还影响柄曲霉素和黄曲霉毒素的产生[70].除了信号分子诱导之外,环境压力胁迫也是次级代谢产物开启的重要因素,例如委内瑞拉链霉菌中杰多霉素B的产生需要噬菌体的入侵或者乙醇的诱导[71].丝状真菌中也存在类似的现象,如黄曲霉中活性氧的产生或者体外添加氧化物能够诱导黄曲霉毒素的产生[72].然而关于群感效应分子和胁迫诱导丝状真菌次级代谢物产生的作用机制尚不清楚,这也是未来丝状真菌次级代谢研究需要加强的地方. ...
Autoregulation of antibiotic biosynthesis by binding of the end product to an atypical response regulator
1
2009
... id="C20">丝状真菌次级代谢物什么时间产生是一个重要的科学问题,普遍认为它产生于生命周期特定的时间点和特定的环境条件下.次级代谢物合成的开启也被认为是初级代谢过程的结束.初级代谢向次级代谢的转换往往与营养耗尽、光照缺少、生殖结构发育、pH变化等密切相关.研究表明构巢曲霉和黄曲霉中次级代谢物合成的开启与有性子实体的形成和菌核的产生有很强的关联性,分生孢子的产生与参与孢子色素合成相关基因的表达也有很强的相关性[69].初级代谢向次级代谢转换的过程中往往伴随着信号分子的产生,而信号分子对次级代谢物的产生具有诱导作用.如群感效应分子在链霉菌次级代谢物合成的开启中起到决定性的作用一样,群感效应分子与丝状真菌的形态分化和次级代谢物的产生也具有相关性.如构巢曲霉和黄曲霉中的群感效应分子氧脂素不仅在调节有性孢子和无性孢子的比例中起关键作用,同时还影响柄曲霉素和黄曲霉毒素的产生[70].除了信号分子诱导之外,环境压力胁迫也是次级代谢产物开启的重要因素,例如委内瑞拉链霉菌中杰多霉素B的产生需要噬菌体的入侵或者乙醇的诱导[71].丝状真菌中也存在类似的现象,如黄曲霉中活性氧的产生或者体外添加氧化物能够诱导黄曲霉毒素的产生[72].然而关于群感效应分子和胁迫诱导丝状真菌次级代谢物产生的作用机制尚不清楚,这也是未来丝状真菌次级代谢研究需要加强的地方. ...
Responses of Aspergillus flavus to oxidative stress are related to fungal development regulator, antioxidant enzyme, and secondary metabolite biosynthetic gene expression
1
2016
... id="C20">丝状真菌次级代谢物什么时间产生是一个重要的科学问题,普遍认为它产生于生命周期特定的时间点和特定的环境条件下.次级代谢物合成的开启也被认为是初级代谢过程的结束.初级代谢向次级代谢的转换往往与营养耗尽、光照缺少、生殖结构发育、pH变化等密切相关.研究表明构巢曲霉和黄曲霉中次级代谢物合成的开启与有性子实体的形成和菌核的产生有很强的关联性,分生孢子的产生与参与孢子色素合成相关基因的表达也有很强的相关性[69].初级代谢向次级代谢转换的过程中往往伴随着信号分子的产生,而信号分子对次级代谢物的产生具有诱导作用.如群感效应分子在链霉菌次级代谢物合成的开启中起到决定性的作用一样,群感效应分子与丝状真菌的形态分化和次级代谢物的产生也具有相关性.如构巢曲霉和黄曲霉中的群感效应分子氧脂素不仅在调节有性孢子和无性孢子的比例中起关键作用,同时还影响柄曲霉素和黄曲霉毒素的产生[70].除了信号分子诱导之外,环境压力胁迫也是次级代谢产物开启的重要因素,例如委内瑞拉链霉菌中杰多霉素B的产生需要噬菌体的入侵或者乙醇的诱导[71].丝状真菌中也存在类似的现象,如黄曲霉中活性氧的产生或者体外添加氧化物能够诱导黄曲霉毒素的产生[72].然而关于群感效应分子和胁迫诱导丝状真菌次级代谢物产生的作用机制尚不清楚,这也是未来丝状真菌次级代谢研究需要加强的地方. ...
The autophagy-related gene Acatg1 is involved in conidiationand cephalosporin production in Acremonium chrysogenum
1
2014
... id="C21">自噬是真核细胞中普遍存在且高度保守的细胞降解和营养再利用过程.而丝状真菌自噬与次级代谢间的关系还鲜有报道.本课题组发现顶头孢霉自噬基因Acatg1、Acatg8和Acatg11均参与孢子的产生和头孢菌素的合成[73, 74].Acatg1和Acatg8正调控分生孢子的产生,但负调控头孢菌素的合成.敲除Acatg1和Acatg8能够通过增强发酵后期头孢菌素合成蛋白的稳定性提高抗生素产量.而Acatg11则负调控分生孢子的产生,但正调控头孢菌素的合成.这说明不同的自噬基因在次级代谢中发挥的功能不同. ...
Functional analysis of the selective autophagy related gene Acatg11 inAcremonium chrysogenum
1
2017
... id="C21">自噬是真核细胞中普遍存在且高度保守的细胞降解和营养再利用过程.而丝状真菌自噬与次级代谢间的关系还鲜有报道.本课题组发现顶头孢霉自噬基因Acatg1、Acatg8和Acatg11均参与孢子的产生和头孢菌素的合成[73, 74].Acatg1和Acatg8正调控分生孢子的产生,但负调控头孢菌素的合成.敲除Acatg1和Acatg8能够通过增强发酵后期头孢菌素合成蛋白的稳定性提高抗生素产量.而Acatg11则负调控分生孢子的产生,但正调控头孢菌素的合成.这说明不同的自噬基因在次级代谢中发挥的功能不同. ...
Genetic manipulation of the COP9 signalosome subunit PfCsnE leads to the discovery of pestaloficins in Pestalotiopsis fici
1
2017
... id="C22">CsnE/Csn5是泛素-蛋白酶体的重要组成部分,并被证明参与丝状真菌形态分化和次级代谢.尹文兵课题组发现敲除PfcsnE导致无花果拟盘多毛孢不再产生分生孢子,同时次级代谢物的合成也受到了影响.PfcsnE突变株中产生了更多的氯代二苯酮,但是ficiolide A的产量却有所下降,同时PfcsnE的敲除还激活了7种化合物的产生[75].转录组分析表明,PfcsnE的存在分别上调了9个以及下调了3个生物合成基因簇的表达[76].浙江大学李红叶课题组敲除Aacsn5导致链格孢(Alternaria alternata)菌丝生长和分生孢子产生都受到了影响,转录组数据表明,Aacsn5调控与分生孢子产生和侵染相关的次级代谢物的合成[77]. ...
COP9 signalosome subunit PfCsnE regulates secondary metabolism and conidial formation in Pestalotiopsis fici
1
2017
... id="C22">CsnE/Csn5是泛素-蛋白酶体的重要组成部分,并被证明参与丝状真菌形态分化和次级代谢.尹文兵课题组发现敲除PfcsnE导致无花果拟盘多毛孢不再产生分生孢子,同时次级代谢物的合成也受到了影响.PfcsnE突变株中产生了更多的氯代二苯酮,但是ficiolide A的产量却有所下降,同时PfcsnE的敲除还激活了7种化合物的产生[75].转录组分析表明,PfcsnE的存在分别上调了9个以及下调了3个生物合成基因簇的表达[76].浙江大学李红叶课题组敲除Aacsn5导致链格孢(Alternaria alternata)菌丝生长和分生孢子产生都受到了影响,转录组数据表明,Aacsn5调控与分生孢子产生和侵染相关的次级代谢物的合成[77]. ...
Csn5 is required for the conidiogenesis and pathogenesis of the Alternaria alternata tangerine pathotype
1
2018
... id="C22">CsnE/Csn5是泛素-蛋白酶体的重要组成部分,并被证明参与丝状真菌形态分化和次级代谢.尹文兵课题组发现敲除PfcsnE导致无花果拟盘多毛孢不再产生分生孢子,同时次级代谢物的合成也受到了影响.PfcsnE突变株中产生了更多的氯代二苯酮,但是ficiolide A的产量却有所下降,同时PfcsnE的敲除还激活了7种化合物的产生[75].转录组分析表明,PfcsnE的存在分别上调了9个以及下调了3个生物合成基因簇的表达[76].浙江大学李红叶课题组敲除Aacsn5导致链格孢(Alternaria alternata)菌丝生长和分生孢子产生都受到了影响,转录组数据表明,Aacsn5调控与分生孢子产生和侵染相关的次级代谢物的合成[77]. ...
The fungal myosin I is essential for Fusarium toxisome formation
1
2018
... id="C23">肌动蛋白Ⅰ抑制剂氰烯菌酯能够特异性抑制肌球蛋白Ⅰ ATP水解酶的活性.马忠华课题组发现,氰烯菌酯通过抑制肌球蛋白Ⅰ ATP水解酶的活性抑制禾谷镰刀菌毒素小体的形成,并降低DON的产生.研究表明肌动蛋白Ⅰ(FgMyo I)还与核糖体相关蛋白FgAsc1互作,调控DON生物合成关键酶Tri1的翻译.然而肌球蛋白Ⅰ与被测试的其他次级代谢物的生物合成不相关.此项研究揭示了肌球蛋白Ⅰ参与调控真菌毒素生物合成的新功能[78].微管末端结合蛋白能够调控酵母细胞微管的动态性、细胞的极性以及染色体的稳定性,而在丝状真菌中的功能不清楚.马忠华课题组发现,微管末端结合蛋白同源基因FgEB1调控禾谷镰刀菌的细胞极性以及对杀菌剂的敏感性,FgEB1缺失显著降低DON产量[79].该课题组还发现禾谷镰刀菌中含有8个几丁质合成酶基因,其中FgChs2、FgChs4、FgChs7和FgChs5的缺失不同程度地削弱了DON的产生,同时FgChs2与其他几丁质酶基因的双缺失进一步降低了DON产量,说明几丁质酶与真菌毒素合成之间存在很强的关联性[80]. ...
The microtubule end-binding protein FgEB1 regulates polar growth and fungicide sensitivity via different interactors in Fusarium graminearum
1
2017
... id="C23">肌动蛋白Ⅰ抑制剂氰烯菌酯能够特异性抑制肌球蛋白Ⅰ ATP水解酶的活性.马忠华课题组发现,氰烯菌酯通过抑制肌球蛋白Ⅰ ATP水解酶的活性抑制禾谷镰刀菌毒素小体的形成,并降低DON的产生.研究表明肌动蛋白Ⅰ(FgMyo I)还与核糖体相关蛋白FgAsc1互作,调控DON生物合成关键酶Tri1的翻译.然而肌球蛋白Ⅰ与被测试的其他次级代谢物的生物合成不相关.此项研究揭示了肌球蛋白Ⅰ参与调控真菌毒素生物合成的新功能[78].微管末端结合蛋白能够调控酵母细胞微管的动态性、细胞的极性以及染色体的稳定性,而在丝状真菌中的功能不清楚.马忠华课题组发现,微管末端结合蛋白同源基因FgEB1调控禾谷镰刀菌的细胞极性以及对杀菌剂的敏感性,FgEB1缺失显著降低DON产量[79].该课题组还发现禾谷镰刀菌中含有8个几丁质合成酶基因,其中FgChs2、FgChs4、FgChs7和FgChs5的缺失不同程度地削弱了DON的产生,同时FgChs2与其他几丁质酶基因的双缺失进一步降低了DON产量,说明几丁质酶与真菌毒素合成之间存在很强的关联性[80]. ...
The chitin synthase FgChs2 and other FgChss co-regulate vegetative development and virulence in F. graminearum
1
2016
... id="C23">肌动蛋白Ⅰ抑制剂氰烯菌酯能够特异性抑制肌球蛋白Ⅰ ATP水解酶的活性.马忠华课题组发现,氰烯菌酯通过抑制肌球蛋白Ⅰ ATP水解酶的活性抑制禾谷镰刀菌毒素小体的形成,并降低DON的产生.研究表明肌动蛋白Ⅰ(FgMyo I)还与核糖体相关蛋白FgAsc1互作,调控DON生物合成关键酶Tri1的翻译.然而肌球蛋白Ⅰ与被测试的其他次级代谢物的生物合成不相关.此项研究揭示了肌球蛋白Ⅰ参与调控真菌毒素生物合成的新功能[78].微管末端结合蛋白能够调控酵母细胞微管的动态性、细胞的极性以及染色体的稳定性,而在丝状真菌中的功能不清楚.马忠华课题组发现,微管末端结合蛋白同源基因FgEB1调控禾谷镰刀菌的细胞极性以及对杀菌剂的敏感性,FgEB1缺失显著降低DON产量[79].该课题组还发现禾谷镰刀菌中含有8个几丁质合成酶基因,其中FgChs2、FgChs4、FgChs7和FgChs5的缺失不同程度地削弱了DON的产生,同时FgChs2与其他几丁质酶基因的双缺失进一步降低了DON产量,说明几丁质酶与真菌毒素合成之间存在很强的关联性[80]. ...
The AMP-activated protein kinase homolog Snf1 concerts carbon utilization, conidia production and the biosynthesis of secondary metabolites in the taxol-producer Pestalotiopsis microspora
1
2018
... id="C24">AMP依赖的蛋白激酶(AMPK)在丝状真菌营养代谢中具有重要作用.朱旭东课题组发现AMPK的同源基因snf1参与小孢拟盘多毛孢的初级代谢、细胞壁合成以及对渗透压的感应,snf1的破坏延迟了气生菌丝的生长,降低了分生孢子数量,同时pestalotiollide B不再产生,这说明Snf1也是参与次级代谢的关键因子[81].陈福生课题组发现G蛋白α亚基编码基因Mga1的缺失导致红曲霉中橘霉素的产量提高了9倍,其他色素的产量也增加了71%,因此Mga1除了参与信号转导途径之外还调控次级代谢物的合成[82].G蛋白信号转导途径中的FlbA同源基因的缺失除了阻碍分生孢子萌发,还抑制了橘霉素和色素的产生[83],蛋白组学的分析也证实了这一结果[84]. ...
Identification of Mga1, a G-protein alpha-subunit gene involved in regulating citrinin and pigment production in Monascus ruber M7
1
2010
... id="C24">AMP依赖的蛋白激酶(AMPK)在丝状真菌营养代谢中具有重要作用.朱旭东课题组发现AMPK的同源基因snf1参与小孢拟盘多毛孢的初级代谢、细胞壁合成以及对渗透压的感应,snf1的破坏延迟了气生菌丝的生长,降低了分生孢子数量,同时pestalotiollide B不再产生,这说明Snf1也是参与次级代谢的关键因子[81].陈福生课题组发现G蛋白α亚基编码基因Mga1的缺失导致红曲霉中橘霉素的产量提高了9倍,其他色素的产量也增加了71%,因此Mga1除了参与信号转导途径之外还调控次级代谢物的合成[82].G蛋白信号转导途径中的FlbA同源基因的缺失除了阻碍分生孢子萌发,还抑制了橘霉素和色素的产生[83],蛋白组学的分析也证实了这一结果[84]. ...
mrflbA, encoding a putative FlbA, is involved in aerial hyphal development and secondary metabolite production in Monascus ruber M-7
1
2012
... id="C24">AMP依赖的蛋白激酶(AMPK)在丝状真菌营养代谢中具有重要作用.朱旭东课题组发现AMPK的同源基因snf1参与小孢拟盘多毛孢的初级代谢、细胞壁合成以及对渗透压的感应,snf1的破坏延迟了气生菌丝的生长,降低了分生孢子数量,同时pestalotiollide B不再产生,这说明Snf1也是参与次级代谢的关键因子[81].陈福生课题组发现G蛋白α亚基编码基因Mga1的缺失导致红曲霉中橘霉素的产量提高了9倍,其他色素的产量也增加了71%,因此Mga1除了参与信号转导途径之外还调控次级代谢物的合成[82].G蛋白信号转导途径中的FlbA同源基因的缺失除了阻碍分生孢子萌发,还抑制了橘霉素和色素的产生[83],蛋白组学的分析也证实了这一结果[84]. ...
Proteome analysis reveals global response to deletion of mrflbA in Monascus ruber
1
2018
... id="C24">AMP依赖的蛋白激酶(AMPK)在丝状真菌营养代谢中具有重要作用.朱旭东课题组发现AMPK的同源基因snf1参与小孢拟盘多毛孢的初级代谢、细胞壁合成以及对渗透压的感应,snf1的破坏延迟了气生菌丝的生长,降低了分生孢子数量,同时pestalotiollide B不再产生,这说明Snf1也是参与次级代谢的关键因子[81].陈福生课题组发现G蛋白α亚基编码基因Mga1的缺失导致红曲霉中橘霉素的产量提高了9倍,其他色素的产量也增加了71%,因此Mga1除了参与信号转导途径之外还调控次级代谢物的合成[82].G蛋白信号转导途径中的FlbA同源基因的缺失除了阻碍分生孢子萌发,还抑制了橘霉素和色素的产生[83],蛋白组学的分析也证实了这一结果[84]. ...
Regulation of development in Aspergillus nidulans and Aspergillus fumigatus
1
2010
... id="C25">真核生物通过外界信号在细胞表面和细胞核之间的传递进行细胞通讯,其中有丝分裂原激活蛋白激酶(MAPK)级联系统是关键因素之一.MAP3K磷酸化MAP2K,MAP2K磷酸化MAPK,而MAPK通过磷酸化细胞核内的目标蛋白从而激活特定的基因表达[85].AnFus3是MAPK在构巢曲霉中的同源蛋白,是负责蛋白入核的重要因子.AnFus3通过与保守的核转录因子AnSte12互作从而起始性发育过程.AnFus3能与磷酸化的VeA互作,进而调控丝状真菌的形态分化和次级代谢[86].这个级联传递系统中的所有因子都参与了粗糙脉孢菌的形态分化、无性和有性发育过程[87].我国学者对MAPK与丝状真菌形态分化和次级代谢间关系的研究较少.哈尔滨商业大学杨萍发现棘孢木霉(T. asperellum)中MAPK基因task1与菌体的形态分化、真菌寄生体的互作以及次级代谢都有很强的相关性.task1的缺失增加了6-戊基-吡喃酮产量,也增强了对病原菌生长的抑制作用[88].河北农业大学董金皋课题组发现玉米大斑病菌(Setosphaeria turcica)中MAPK编码基因StPBS2参与菌体的形态分化、分生孢子产生、细胞壁发育、耐药性以及次级代谢物的合成.StPBS2受到干扰的菌株与野生株相比产生了更多红色的次级代谢物,而且基因干扰的程度与次级代谢物的量呈正相关[89]. ...
The Aspergillus nidulans MAPK module AnSte11-Ste50-Ste7- Fus3 controls development and secondary metabolism
1
2012
... id="C25">真核生物通过外界信号在细胞表面和细胞核之间的传递进行细胞通讯,其中有丝分裂原激活蛋白激酶(MAPK)级联系统是关键因素之一.MAP3K磷酸化MAP2K,MAP2K磷酸化MAPK,而MAPK通过磷酸化细胞核内的目标蛋白从而激活特定的基因表达[85].AnFus3是MAPK在构巢曲霉中的同源蛋白,是负责蛋白入核的重要因子.AnFus3通过与保守的核转录因子AnSte12互作从而起始性发育过程.AnFus3能与磷酸化的VeA互作,进而调控丝状真菌的形态分化和次级代谢[86].这个级联传递系统中的所有因子都参与了粗糙脉孢菌的形态分化、无性和有性发育过程[87].我国学者对MAPK与丝状真菌形态分化和次级代谢间关系的研究较少.哈尔滨商业大学杨萍发现棘孢木霉(T. asperellum)中MAPK基因task1与菌体的形态分化、真菌寄生体的互作以及次级代谢都有很强的相关性.task1的缺失增加了6-戊基-吡喃酮产量,也增强了对病原菌生长的抑制作用[88].河北农业大学董金皋课题组发现玉米大斑病菌(Setosphaeria turcica)中MAPK编码基因StPBS2参与菌体的形态分化、分生孢子产生、细胞壁发育、耐药性以及次级代谢物的合成.StPBS2受到干扰的菌株与野生株相比产生了更多红色的次级代谢物,而且基因干扰的程度与次级代谢物的量呈正相关[89]. ...
Mitogen-activated protein kinase cascade required for regulation of development and secondary metabolism in Neurospora crassa
1
2008
... id="C25">真核生物通过外界信号在细胞表面和细胞核之间的传递进行细胞通讯,其中有丝分裂原激活蛋白激酶(MAPK)级联系统是关键因素之一.MAP3K磷酸化MAP2K,MAP2K磷酸化MAPK,而MAPK通过磷酸化细胞核内的目标蛋白从而激活特定的基因表达[85].AnFus3是MAPK在构巢曲霉中的同源蛋白,是负责蛋白入核的重要因子.AnFus3通过与保守的核转录因子AnSte12互作从而起始性发育过程.AnFus3能与磷酸化的VeA互作,进而调控丝状真菌的形态分化和次级代谢[86].这个级联传递系统中的所有因子都参与了粗糙脉孢菌的形态分化、无性和有性发育过程[87].我国学者对MAPK与丝状真菌形态分化和次级代谢间关系的研究较少.哈尔滨商业大学杨萍发现棘孢木霉(T. asperellum)中MAPK基因task1与菌体的形态分化、真菌寄生体的互作以及次级代谢都有很强的相关性.task1的缺失增加了6-戊基-吡喃酮产量,也增强了对病原菌生长的抑制作用[88].河北农业大学董金皋课题组发现玉米大斑病菌(Setosphaeria turcica)中MAPK编码基因StPBS2参与菌体的形态分化、分生孢子产生、细胞壁发育、耐药性以及次级代谢物的合成.StPBS2受到干扰的菌株与野生株相比产生了更多红色的次级代谢物,而且基因干扰的程度与次级代谢物的量呈正相关[89]. ...
The gene task1 is involved in morphological development, mycoparasitism and antibiosis of Trichoderma asperellum
1
2017
... id="C25">真核生物通过外界信号在细胞表面和细胞核之间的传递进行细胞通讯,其中有丝分裂原激活蛋白激酶(MAPK)级联系统是关键因素之一.MAP3K磷酸化MAP2K,MAP2K磷酸化MAPK,而MAPK通过磷酸化细胞核内的目标蛋白从而激活特定的基因表达[85].AnFus3是MAPK在构巢曲霉中的同源蛋白,是负责蛋白入核的重要因子.AnFus3通过与保守的核转录因子AnSte12互作从而起始性发育过程.AnFus3能与磷酸化的VeA互作,进而调控丝状真菌的形态分化和次级代谢[86].这个级联传递系统中的所有因子都参与了粗糙脉孢菌的形态分化、无性和有性发育过程[87].我国学者对MAPK与丝状真菌形态分化和次级代谢间关系的研究较少.哈尔滨商业大学杨萍发现棘孢木霉(T. asperellum)中MAPK基因task1与菌体的形态分化、真菌寄生体的互作以及次级代谢都有很强的相关性.task1的缺失增加了6-戊基-吡喃酮产量,也增强了对病原菌生长的抑制作用[88].河北农业大学董金皋课题组发现玉米大斑病菌(Setosphaeria turcica)中MAPK编码基因StPBS2参与菌体的形态分化、分生孢子产生、细胞壁发育、耐药性以及次级代谢物的合成.StPBS2受到干扰的菌株与野生株相比产生了更多红色的次级代谢物,而且基因干扰的程度与次级代谢物的量呈正相关[89]. ...
StPBS2, a MAPK kinase gene, is involved in determining hyphal morphology, cell wall development, hypertonic stress reaction as well as the production of secondary metabolites in Northern Corn Leaf Blight pathogen Setosphaeria turcica
1
2017
... id="C25">真核生物通过外界信号在细胞表面和细胞核之间的传递进行细胞通讯,其中有丝分裂原激活蛋白激酶(MAPK)级联系统是关键因素之一.MAP3K磷酸化MAP2K,MAP2K磷酸化MAPK,而MAPK通过磷酸化细胞核内的目标蛋白从而激活特定的基因表达[85].AnFus3是MAPK在构巢曲霉中的同源蛋白,是负责蛋白入核的重要因子.AnFus3通过与保守的核转录因子AnSte12互作从而起始性发育过程.AnFus3能与磷酸化的VeA互作,进而调控丝状真菌的形态分化和次级代谢[86].这个级联传递系统中的所有因子都参与了粗糙脉孢菌的形态分化、无性和有性发育过程[87].我国学者对MAPK与丝状真菌形态分化和次级代谢间关系的研究较少.哈尔滨商业大学杨萍发现棘孢木霉(T. asperellum)中MAPK基因task1与菌体的形态分化、真菌寄生体的互作以及次级代谢都有很强的相关性.task1的缺失增加了6-戊基-吡喃酮产量,也增强了对病原菌生长的抑制作用[88].河北农业大学董金皋课题组发现玉米大斑病菌(Setosphaeria turcica)中MAPK编码基因StPBS2参与菌体的形态分化、分生孢子产生、细胞壁发育、耐药性以及次级代谢物的合成.StPBS2受到干扰的菌株与野生株相比产生了更多红色的次级代谢物,而且基因干扰的程度与次级代谢物的量呈正相关[89]. ...



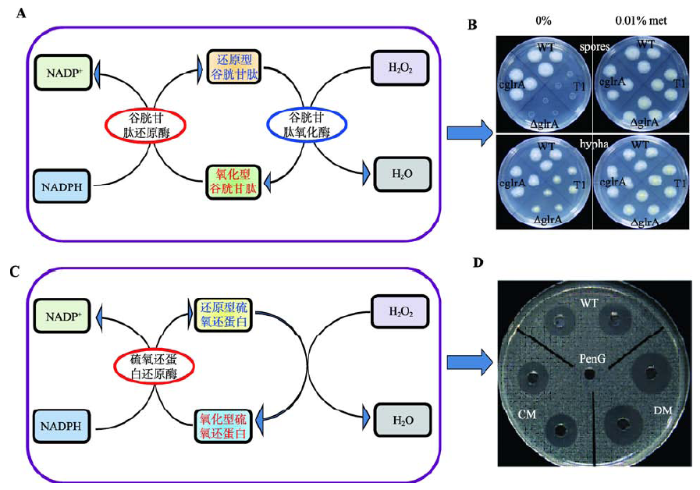
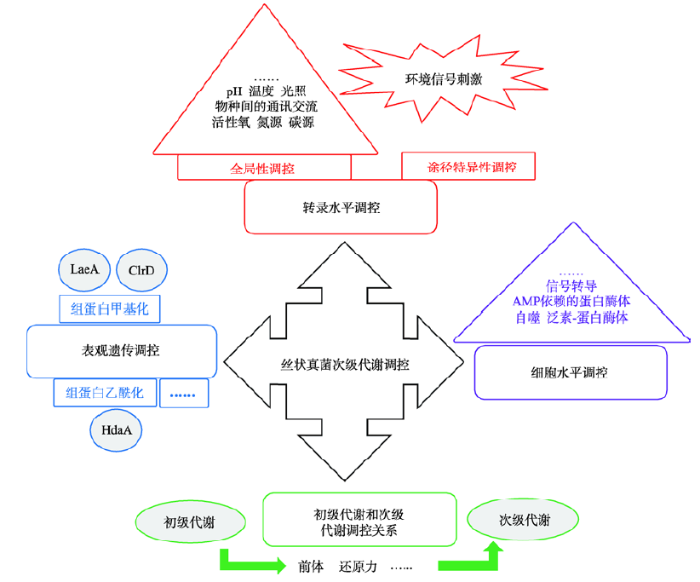
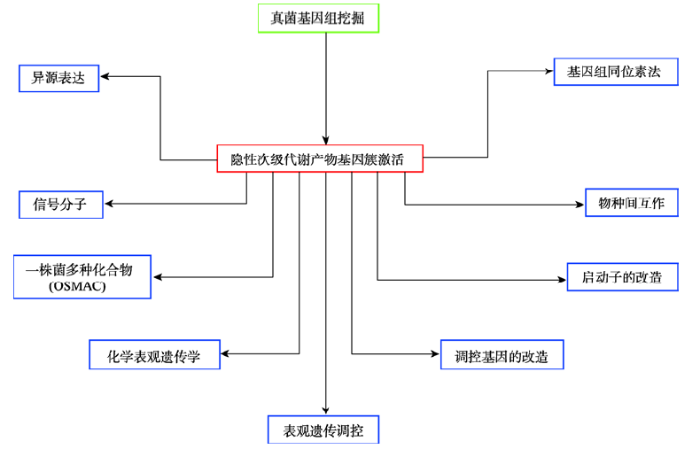




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}